Cấu Trúc Tế Bào Thực Vật. Mô Thực Vật. Các Mô Thực Vật Dẫn điện ...
Có thể bạn quan tâm
Trong sinh học, mô là một nhóm tế bào có cấu trúc và nguồn gốc giống nhau, đồng thời cũng thực hiện các chức năng giống nhau. Ở thực vật, các mô đa dạng và phức tạp nhất đã phát triển trong quá trình tiến hóa ở thực vật hạt kín (thực vật có hoa). Các cơ quan thực vật thường được hình thành bởi một số mô. Sáu loại mô thực vật có thể được phân biệt: giáo dục, cơ bản, dẫn điện, cơ học, nguyên sinh, tiết. Mỗi loại vải bao gồm các loại phụ. Giữa các mô, cũng như bên trong chúng, có khoảng gian bào - khoảng trống giữa các tế bào.
Vải giáo dục
Thông qua phân chia tế bào mô giáo dục cây phát triển theo chiều dài và độ dày. Trong trường hợp này, một phần tế bào của mô giáo dục biệt hóa thành tế bào của các mô khác.
Các tế bào của mô giáo dục đủ nhỏ, liền kề chặt chẽ với nhau, có nhân lớn và vỏ mỏng.
Mô giáo dục ở thực vật ở nón tăng trưởng rễ (đầu rễ) và thân (đỉnh thân), nó xảy ra ở các gốc của các lóng, còn mô giáo dục là cambium(đảm bảo sự phát triển của thân cây theo chiều dày).
Tế bào nón sinh trưởng của rễ. Ảnh chụp quá trình phân bào (phân kì của nhiễm sắc thể, phân li của nhân).
Nhu mô, hoặc mô chính
Nhu mô bao gồm một số loại mô. Phân biệt giữa đồng hóa (quang hợp), lưu trữ, tầng chứa nước và mô cơ bản trong không khí.
Mô quang hợp bao gồm các tế bào chứa chất diệp lục, tức là các tế bào màu xanh lục. Những tế bào này có thành mỏng và chứa một số lượng lớn lục lạp. Chức năng chính của chúng là quang hợp. Mô đồng hóa cấu tạo nên cùi của lá, là phần vỏ của thân cây non và thân cỏ.

Trong lồng mô lưu trữ cổ phiếu đang tích lũy chất dinh dưỡng... Mô này tạo nên nội nhũ của hạt, là một phần của củ, củ, ... Phần lõi của thân, các tế bào bên trong vỏ của thân và rễ, lớp vỏ mọng nước cũng thường bao gồm một nhu mô chứa.
Nhu mô mọng nước chỉ đặc trưng của một số loài thực vật, thường là nơi sống khô cằn. Nước tích tụ trong các tế bào của mô này. Mô chứa nước có thể được tìm thấy cả trong lá (lô hội) và thân (xương rồng).
Khăn giấy thoáng khíđặc trưng của thực vật thủy sinh và đầm lầy. Đặc điểm của nó là sự hiện diện của một số lượng lớn các khoảng gian bào chứa không khí. Điều này giúp cây dễ dàng trao đổi khí khi gặp khó khăn.
Vải dẫn điện
Chức năng chung của các mô dẫn điện khác nhau là mang các chất từ cơ quan thực vật này sang cơ quan thực vật khác. Ở các thân của cây thân gỗ, các tế bào của mô dẫn nằm trong gỗ và libe. Hơn nữa, trong gỗ có vị trí mạch (khí quản) và khí quản, cùng với đó dung dịch nước di chuyển từ rễ, và trong lớp đệm - ống sàng, theo đó chất hữu cơ từ lá cây quang hợp di chuyển.
Mạch và khí quản là những tế bào chết. Dung dịch nước đi qua các mạch nhanh hơn qua các khí quản.
Các ống rây là những tế bào sống, nhưng không có nhân.
Phủ khăn giấy
Mô liên kết bao gồm da (biểu bì), lớp bần, lớp vỏ. Da bao phủ lá và thân xanh, đây là những tế bào sống. Nút chai bao gồm Tế bào chếtđược tẩm một chất giống như chất béo không cho nước và không khí đi qua.
Chức năng chính của bất kỳ mô liên kết nào là bảo vệ tế bào bên trong cây trồng khỏi bị hư hại cơ học, khô héo, sự xâm nhập của vi sinh vật, thay đổi nhiệt độ.
Cork là một mô nguyên sinh thứ cấp, vì nó xuất hiện ở vị trí vỏ của thân và rễ cây lâu năm.
Lớp vỏ bao gồm lớp bần và lớp mô chết bên dưới.
Vải cơ khí
Các tế bào của mô cơ học được đặc trưng bởi các màng hóa lỏng dày lên rất mạnh. Chức năng của mô cơ học là truyền sức mạnh và tính đàn hồi cho cơ thể và các cơ quan của thực vật.
Trong thân của thực vật hạt kín, mô cơ học có thể nằm trong một lớp nguyên vẹn hoặc trong các sợi riêng biệt cách xa nhau.
Ở lá, các sợi của mô cơ thường nằm cạnh các sợi của mô dẫn. Chúng cùng nhau tạo thành các đường gân của lá.
Mô tiết hoặc mô bài tiết của thực vật
Tế bào mô tiết tiết ra các chất khác nhau, và do đó các chức năng của loại vải này khác nhau. Tế bào bài tiết ở thực vật tạo đường dẫn nhựa và tinh dầu, hình thành các tuyến đặc biệt và lông tuyến. Mô tiết gồm các ống hoa.
Nhựa thực hiện chức năng bảo vệ khi thân cây bị hại.
Mật hoa thu hút côn trùng thụ phấn.
Có những tế bào bài tiết các sản phẩm trao đổi chất, ví dụ, muối của axit oxalic.
6.1. Giá trị và sự đa dạng của các mô dẫn điện
Mô dẫn điện là một phần thiết yếu của hầu hết các thực vật bậc cao... Chúng là thành phần cấu tạo không thể thiếu của cơ quan sinh dưỡng và sinh sản của thực vật bào tử và thực vật có hạt. Các mô dẫn, cùng với thành tế bào và khoảng gian bào, một số tế bào của nhu mô chính và các tế bào dẫn truyền chuyên biệt, tạo thành một hệ thống dẫn cung cấp sự vận chuyển các chất theo hướng xa và xuyên tâm. Do cấu trúc đặc biệt của tế bào và vị trí của chúng trong cơ thể thực vật, hệ thống dẫn điện thực hiện nhiều chức năng nhưng có liên quan với nhau:
1) chuyển động của nước và chất khoángđược rễ hấp thụ từ đất, cũng như chất hữu cơ hình thành ở rễ, ở thân, lá, cơ quan sinh sản;
2) sự di chuyển của các sản phẩm quang hợp từ các bộ phận xanh của cây đến nơi sử dụng và bảo quản: ở rễ, thân, quả và hạt;
3) sự di chuyển của phytohormon trong thực vật, tạo ra sự cân bằng nhất định của chúng, quyết định tốc độ sinh trưởng và phát triển của các cơ quan sinh dưỡng và sinh sản của thực vật;
4) sự vận chuyển xuyên tâm của các chất từ các mô dẫn đến các tế bào sống lân cận của các mô khác, ví dụ, để đồng hóa các tế bào của trung bì lá và các tế bào phân chia của mô phân sinh. Các tế bào nhu mô của tia gốc của gỗ và vỏ cây cũng có thể tham gia vào đó. Tầm quan trọng lớn trong vận chuyển xuyên tâm, chúng có các tế bào chuyển với nhiều chỗ lồi của màng tế bào, nằm giữa mô dẫn và nhu mô;
5) các mô dẫn điện làm tăng sức đề kháng của các cơ quan thực vật đối với tải trọng biến dạng;
6) các mô dẫn tạo thành một hệ thống phân nhánh liên tục kết nối các cơ quan thực vật thành một tổng thể duy nhất;
Sự xuất hiện của các mô dẫn là kết quả của quá trình biến đổi cấu trúc tiến hóa liên quan đến sự xuất hiện của thực vật trên cạn và sự tách biệt của không khí và dinh dưỡng trong đất của chúng. Các mô dẫn truyền cổ xưa nhất, khí quản, được tìm thấy trong các hóa thạch tê giác. Chúng đạt đến sự phát triển cao nhất trong thực vật hạt kín hiện đại.
Trong quá trình phát triển cá thể, các mô dẫn sơ cấp được hình thành từ procambium tại các điểm phát triển của phôi hạt và các chồi của sự đổi mới. Các mô dẫn điện thứ cấp, đặc trưng của thực vật hạt kín hai lá mầm, được tạo ra bởi cambium.
Tùy thuộc vào các chức năng được thực hiện, các mô dẫn điện được chia thành các mô của dòng điện tăng dần và các mô của dòng điện giảm dần. Mục đích chính của các mô của dòng điện tăng dần là vận chuyển nước và các chất khoáng hòa tan trong nó từ rễ đến các cơ quan nằm trên mặt đất. Ngoài ra, các chất hữu cơ hình thành trong rễ và thân di chuyển dọc theo chúng, ví dụ như axit hữu cơ, carbohydrate và phytohormone. Tuy nhiên, thuật ngữ "dòng điện đi lên" không nên được hiểu rõ ràng là chuyển động từ dưới lên trên. Các mô của dòng điện tăng dần cung cấp dòng chảy của các chất theo hướng từ vùng hút đến đỉnh chồi. Trong trường hợp này, các chất được vận chuyển được sử dụng cho cả bản thân rễ và thân, cành, lá, cơ quan sinh sản, bất kể chúng ở trên hay dưới mức của rễ. Ví dụ, trong khoai tây, nước và các nguyên tố dinh dưỡng khoáng đi qua các mô của dòng điện tăng dần đến các thân và củ được hình thành trong đất, cũng như đến các cơ quan trên mặt đất.
Các mô Downdraft đảm bảo dòng chảy của các sản phẩm quang hợp vào các bộ phận đang phát triển của thực vật và vào các cơ quan dự trữ. Trong trường hợp này, vị trí không gian của các cơ quan quang hợp không quan trọng. Ví dụ, trong lúa mì, chất hữu cơ xâm nhập vào tế bào ký sinh đang phát triển từ các lá của các tầng khác nhau. Do đó, tên gọi các loại vải “tăng dần” và “giảm dần” không nên được coi là một truyền thống lâu đời.
6.2. Mô dẫn điện tăng dần
Các mô của dòng điện tăng dần bao gồm khí quản và mạch (khí quản), nằm trong phần thân gỗ (xylem) của các cơ quan thực vật. Trong các mô này, sự chuyển động của nước và các chất hòa tan trong nó diễn ra thụ động dưới tác dụng của áp suất rễ và sự bốc hơi nước từ bề mặt của cây.
Tracheids có nguồn gốc cổ xưa hơn. Chúng được tìm thấy ở thực vật bào tử bậc cao, thực vật hạt trần, và ít thường xuyên hơn ở thực vật hạt kín. Ở thực vật hạt kín, chúng đặc trưng cho sự phân nhánh nhỏ nhất của gân lá. Các tế bào khí quản đã chết. Chúng có hình dạng thuôn dài, thường có hình dạng mờ. Chiều dài của chúng là 1 - 4 mm. Tuy nhiên, ở cây hạt trần, ví dụ, ở araucaria, nó đạt tới 10 mm. Thành tế bào dày, có chất xenlulo, thường được tẩm lignin. Màng tế bào chứa nhiều lỗ chân lông có viền.
Các tàu được hình thành ở các giai đoạn sau của quá trình tiến hóa. Chúng là đặc điểm của thực vật hạt kín, mặc dù chúng cũng được tìm thấy ở một số đại diện hiện đại của các phân ngành Chân quỳ (chi Sellaginella), Đuôi ngựa, Dương xỉ và Thực vật hạt trần (chi Gnetum).
Mạch được tạo thành từ các tế bào chết kéo dài, cái này ở trên cái kia, được gọi là các đoạn mạch. Ở thành cuối của các phân đoạn của bình có các lỗ thông lớn - lỗ thủng, qua đó vận chuyển các chất đi đường dài được thực hiện. Các lỗ thủng đã phát sinh trong quá trình tiến hóa từ các lỗ giáp ranh của khí quản. Trong thành phần của các bình, chúng có dạng bậc thang và đơn giản. Nhiều lỗ thủng có tỷ lệ được hình thành trên thành cuối của các phân đoạn của bình khi chúng được đặt xiên. Các lỗ của các lỗ đục như vậy có hình dạng kéo dài và các vách ngăn ngăn cách chúng nằm song song với nhau, giống như các bậc của cầu thang. Vòi có lỗ thủng bậc thang là điển hình cho các cây thuộc họ Bơ, Sả, Bạch dương, Cọ, Chastukhovye.
Các lỗ đơn giản được biết đến trong các họ tiến hóa trẻ hơn, chẳng hạn như Solanaceae, Pumpkin, Aster, Bluegrass. Chúng đại diện cho một lỗ lớn ở thành cuối của phân đoạn, nằm vuông góc với trục của tàu. Ví dụ, trong một số họ, trong số các họ Magnolia, Rose, Iris, Astrov, cả lỗ đục đơn giản và lỗ bậc thang đều được tìm thấy trong các bình.
Các thành bên có độ dày xenluloza không đồng đều giúp bảo vệ mạch khỏi áp suất dư thừa do các tế bào sống lân cận của các mô khác tạo ra. Có thể có nhiều lỗ rỗng ở thành bên để nước thoát ra khỏi bình.
Tùy thuộc vào bản chất của sự dày lên, các loại và bản chất của vị trí của các lỗ chân lông, các mạch máu được chia nhỏ thành hình khuyên, hình xoắn ốc, hình đôi, lưới, hình thang và lỗ chân lông điểm. Trong các mạch hình khuyên và mạch xoắn ốc, các chất dày xenluloza được sắp xếp theo dạng vòng hoặc hình xoắn ốc. Thông qua các khu vực không dày đặc, các dung dịch được vận chuyển sẽ được khuếch tán vào các mô xung quanh. Đường kính của các bình này tương đối nhỏ. Trong các mạch lỗ chân lông hình lưới, có vảy và lỗ thủng, toàn bộ thành bên, ngoại trừ các vị trí của lỗ chân lông đơn giản, dày lên và thường được ngâm tẩm với lignin. Do đó, sự vận chuyển xuyên tâm của các chất trong chúng được thực hiện thông qua nhiều lỗ rỗng dài và chính xác.
Tàu có thời hạn sử dụng. Chúng có thể bị phá hủy do sự tắc nghẽn của các tế bào nhu mô lân cận, cũng như dưới tác động của lực hướng tâm của áp lực các tế bào gỗ mới được hình thành bởi cambium. Trong quá trình tiến hóa, các con tàu trải qua những thay đổi. Các đoạn mạch trở nên ngắn hơn và dày hơn, các vách ngăn ngang xiên được thay thế bằng các đoạn thẳng, và các lỗ đục bậc thang cũng đơn giản.
6.3. Mô dẫn điện Downdraft
Các mô hạ nguồn bao gồm các tế bào sàng và ống sàng với các tế bào đồng hành. Tế bào rây có nguồn gốc cổ xưa hơn. Chúng được tìm thấy ở thực vật bào tử bậc cao và cây hạt trần. Chúng là những tế bào sống, thuôn dài với đầu nhọn. Ở trạng thái trưởng thành, chúng chứa các hạt nhân như một phần của nguyên sinh chất. Trong thành bên của chúng, tại các điểm tiếp xúc của các tế bào liền kề, có các lỗ nhỏ xuyên qua, được gom lại thành từng nhóm và tạo thành các trường sàng mà các chất di chuyển qua đó.
Ống sàng được cấu tạo bởi hàng dọc tế bào kéo dài, ngăn cách bởi vách ngang và được gọi là tấm sàng, trong đó có các trường sàng. Nếu một tấm sàng có một trường sàng thì nó được coi là đơn giản, và nếu nhiều tấm sàng thì phức tạp. Các trường sàng được hình thành bởi nhiều lỗ thông qua - lỗ sàng có đường kính nhỏ. Plasmodesmata đi qua các lỗ này từ tế bào này sang tế bào khác. Polysaccharide callose được đặt trên thành của các lỗ thủng, làm giảm lumen của các lỗ thủng. Khi ống sàng già đi, callose hoàn toàn làm tắc các lỗ và ống ngừng hoạt động.
Trong quá trình hình thành ống rây, một loại protein phloem đặc biệt (protein F) được tổng hợp trong các tế bào tạo thành chúng, và một không bào lớn phát triển. Nó đẩy tế bào chất và nhân về phía vách tế bào. Sau đó, màng không bào bị phá hủy và không gian bên trong của tế bào chứa đầy hỗn hợp tế bào chất và dịch tế bào. Các cơ thể F-protein mất đường viền riêng biệt, hợp nhất lại, tạo thành các sợi gần các tấm rây. Các sợi của chúng đi qua các lỗ thủng từ đoạn này sang đoạn khác của ống sàng. Một hoặc hai tế bào đồng hành gắn chặt vào các đoạn của ống rây, có hình dạng thuôn dài, thành mỏng và tế bào chất sống có nhân và nhiều ti thể. Trong ti thể, ATP được tổng hợp, cần thiết cho sự vận chuyển các chất qua các ống rây. Trong thành của các tế bào đồng hành, có một số lượng lớn các lỗ có plasmasdesmata, gần như gấp 10 lần số lượng của chúng trong các tế bào khác của trung bì lá. Bề mặt của nguyên sinh chất của các tế bào này tăng lên đáng kể do có nhiều nếp gấp được tạo thành bởi plasmalemma.
Tốc độ di chuyển của các chất đồng hóa qua các ống rây vượt quá tốc độ khuếch tán tự do của các chất một cách đáng kể và đạt 50 - 150 cm / h, điều này cho thấy sự vận chuyển tích cực của các chất sử dụng năng lượng của ATP.
Thời gian hoạt động của ống rây ở cây hai lá mầm lâu năm là 1 - 2 năm. Để thay thế chúng, cambium liên tục hình thành các phần tử dẫn điện mới. Ở những con đơn tính thiếu cambium, các ống sàng tồn tại lâu hơn.
6.4. Dẫn dầm
Các mô dẫn nằm trong các cơ quan thực vật dưới dạng sợi dọc, tạo thành bó dẫn. Có bốn loại bó mạch: đơn giản, chung, phức tạp và bó mạch xơ.
Các bó đơn giản được cấu tạo bởi một loại mô dẫn. Ví dụ, ở phần rìa của phiến lá của nhiều loài thực vật, có các bó mạch và khí quản đường kính nhỏ, và ở các chồi có hoa của họ Hoa môi, chỉ từ các ống rây.
Các bó chung được hình thành bởi khí quản, mạch và ống rây. Đôi khi thuật ngữ này được sử dụng để chỉ các bó metame đi qua các lóng và là dấu vết của lá. Các bó phức tạp bao gồm các mô dẫn và nhu mô. Hoàn hảo nhất, đa dạng về cấu trúc và vị trí là các bó mạch dạng sợi.
Các bó sợi có mạch là đặc trưng của nhiều loài thực vật bậc cao và cây hạt trần. Tuy nhiên, chúng là điển hình nhất của thực vật hạt kín. Trong các gói như vậy, các phần khác nhau về chức năng được phân biệt - phloem và xylem. Phloem đảm bảo dòng chảy của các chất đồng hóa từ lá và sự di chuyển của chúng đến nơi sử dụng hoặc lưu trữ. Thông qua xylem, nước và các chất hòa tan trong nó di chuyển từ hệ thống rễ đến lá và các cơ quan khác. Thể tích của phần xylem lớn hơn nhiều lần so với thể tích của phần phloem, vì thể tích nước đi vào cây vượt quá thể tích của các chất đồng hóa được hình thành, do một phần đáng kể nước được cây bay hơi.
Sự đa dạng của bó sợi mạch được xác định bởi nguồn gốc, thành phần mô học và vị trí của chúng trong cây. Nếu các bó được hình thành từ procambium và hoàn thành quá trình phát triển của chúng khi nguồn cung cấp các tế bào của mô giáo dục được sử dụng, như trong đơn phân, chúng được gọi là khép kín để tăng trưởng. Ngược lại, trong chậu dicots, các bó mở không bị giới hạn về tốc độ tăng trưởng, vì chúng được hình thành bởi một cambium và tăng đường kính trong suốt thời gian tồn tại của cây. Ngoài các bó dẫn điện, các bó sợi mạch máu có thể bao gồm các bó mạch chính và vải cơ khí... Ví dụ, ở cây hai lá mầm, phloem được hình thành bởi các ống rây (mô dẫn tăng dần), nhu mô libe (mô chính), và các sợi libe (mô cơ). Xylem bao gồm các mạch và khí quản (mô dẫn xuống), nhu mô thân gỗ (mô chính) và sợi gỗ (mô cơ). Thành phần mô học của xylem và phloem được xác định về mặt di truyền và có thể được sử dụng trong phân loại thực vật để chẩn đoán các đơn vị phân loại khác nhau. Ngoài ra, mức độ phát triển các bộ phận thành phần chùm có thể thay đổi dưới ảnh hưởng của các điều kiện phát triển của thực vật.
Một số loại bó sợi mạch máu đã được biết đến.
Bó mạch khép kín là đặc điểm của lá và thân của thực vật hạt kín một lá mầm. Họ thiếu cambium. Phloem và xylem được đặt cạnh nhau. Chúng được đặc trưng bởi một số tính năng thiết kế... Vì vậy, ở lúa mì, khác với con đường C 3 của quá trình quang hợp, các bó được hình thành từ procambium và có một phloem sơ cấp và xylem sơ cấp. Trong phloem, một protofloem sớm hơn và muộn hơn trong thời gian hình thành, nhưng một siêu tế bào lớn hơn, được phân biệt. Phần phloem thiếu nhu mô và sợi libe. Trong xylem, ban đầu, các mạch nhỏ hơn của protoxylem được hình thành, nằm trên một đường vuông góc với đường viền bên trong của phloem. Metaxylem được đại diện bởi hai mạch lớn nằm cạnh metafloem vuông góc với chuỗi mạch protoxylem. Trong trường hợp này, các bình được sắp xếp theo hình chữ T. Sự sắp xếp các mạch hình chữ V, Y- và È cũng được biết đến. Giữa các mạch của metaxylem xếp thành 1 - 2 hàng, có một mô xơ cứng tế bào nhỏ với thành dày, được tẩm chất lignin khi thân phát triển. Sclerenchyma này tách vùng xylem ra khỏi phloem. Ở cả hai mặt của mạch của tế bào thân, các tế bào của nhu mô thân gỗ, có thể đóng vai trò truyền máu, vì trong quá trình chuyển bó từ lóng sang đệm lá của nút thân, chúng tham gia vào quá trình hình thành. của tế bào truyền dẫn. Xung quanh bó dẫn của thân lúa mì là lớp vỏ mô cứng, được phát triển tốt hơn từ phía của protoxylem và protofloem; gần các mặt bên của bó, các tế bào của bẹ xếp thành một hàng.
Ở thực vật có kiểu quang hợp C 4 (ngô, kê, v.v.), một bẹ chứa nhiều tế bào chlorenchyme lớn nằm trong lá xung quanh các bó mạch kín.
Các bó nhánh hở là đặc điểm của thân cây hai lá mầm. Sự hiện diện của một lớp cambium giữa phloem và xylem, cũng như không có lớp vỏ mô cứng xung quanh các bó, đảm bảo sự phát triển lâu dài về độ dày của chúng. Trong các phần xylem và phloem của các bó như vậy, có các tế bào của mô chính và cơ.
Các gói tài sản thế chấp mở có thể được hình thành theo hai cách. Thứ nhất, đây là những bó chủ yếu được hình thành bởi procambium. Sau đó, cambium phát triển trong chúng từ các tế bào của nhu mô chính, tạo ra các yếu tố phụ của phloem và xylem. Kết quả là, các chùm sẽ kết hợp các yếu tố mô học có nguồn gốc chính và phụ. Những chùm như vậy là đặc điểm của nhiều loài thực vật có hoa thân thảo thuộc lớp Hai lá mầm, có kiểu cấu trúc thân chùm (họ đậu, họ hoa hồng, v.v.).
Thứ hai, các bó thế chấp mở chỉ có thể được hình thành bởi cambium và bao gồm xylem và phloem có nguồn gốc thứ cấp. Chúng điển hình cho cây hai lá mầm thân thảo với kiểu chuyển tiếp cấu trúc giải phẫu của thân (củ mài, v.v.), cũng như cho cây lấy củ như củ cải đường.
Ở thân cây của một số họ (họ Bí đỏ, họ Solanaceae, họ Kolokolchikovye, v.v.), có các bó hở hai bên, ở đó các xylem được bao bọc bởi các phloem ở cả hai mặt. Trong trường hợp này, phần bên ngoài của phloem đối diện với bề mặt của thân cây phát triển tốt hơn phần bên trong và dải cambi, theo quy luật, nằm giữa xylem và phần ngoài của phloem.
Dầm đồng tâm có hai loại. Trong các bó thân rễ, điển hình của thân rễ dương xỉ, các phloem bao quanh xylem, trong các bó amphivasal, các xylem nằm trong một vòng xung quanh các phloem (thân rễ của mống mắt, hoa huệ thung lũng, v.v.). Ít thường xuyên hơn, các bó đồng tâm được tìm thấy ở cây hai lá mầm (cây thầu dầu).
Các bó mạch kín hướng tâm được hình thành ở những vùng của rễ có cấu trúc giải phẫu sơ cấp. Bó hướng tâm là một phần của hình trụ trung tâm và đi qua phần giữa của rễ. Xylem của nó trông giống như một ngôi sao nhiều tia. Tế bào phloem nằm giữa các tia xylem. Số lượng tia xylem phần lớn phụ thuộc vào bản chất di truyền của cây. Ví dụ, ở cà rốt, củ cải đường, bắp cải và các cây hai lá mầm khác, xylem của bó hướng tâm chỉ có hai tia. Một cây táo và một quả lê có thể có 3 - 5 quả, bí ngô và đậu có xylem 4 tia, và cây đơn tính có một cây nhiều tia. Sự sắp xếp xuyên tâm của các tia xylem là thích nghi. Nó rút ngắn đường đi của nước từ bề mặt hút của rễ đến các mạch của trụ trung tâm.
Ở các cây thân gỗ lâu năm và một số cây thân thảo, ví dụ như ở cây lanh, các mô dẫn nằm trong thân cây mà không hình thành các bó dẫn riêng biệt. Sau đó, họ nói về một loại cấu trúc thân cây không bó.
6.5. Mô vận chuyển xuyên tâm
Các mô cụ thể điều chỉnh sự vận chuyển xuyên tâm của các chất bao gồm ngoại bì và nội bì.
Ngoại bì là lớp ngoài cùng của vỏ rễ sơ cấp. Nó được hình thành ngay dưới lớp mô nguyên sinh nguyên sinh ở vùng lông rễ và bao gồm một hoặc nhiều lớp tế bào đóng chặt với màng xenluloza dày. Trong ngoại bì, nước xâm nhập vào rễ qua các lông hút của rễ sẽ bị tế bào chất nhớt kháng lại và di chuyển vào màng xenlulo của tế bào ngoại bì, sau đó đưa chúng vào các khoảng gian bào của lớp giữa của vỏ sơ cấp hay còn gọi là trung bì. Điều này đảm bảo rằng nước chảy hiệu quả vào các lớp sâu hơn của rễ.
Trong vùng dẫn truyền ở rễ đơn tính, nơi các tế bào của biểu bì chết đi và bong ra, biểu bì xuất hiện trên bề mặt của rễ. Thành tế bào của nó được tẩm suberin và ngăn dòng nước từ đất đến rễ. Ở cây hai lá mầm, lớp vỏ ngoài ở vỏ sơ cấp bong ra trong quá trình lột xác của rễ và được thay thế bằng lớp vỏ ngoài.
Nội bì, hay lớp trong của vỏ rễ sơ cấp, nằm xung quanh hình trụ trung tâm. Nó được hình thành bởi một lớp tế bào đóng chặt có cấu trúc không bằng nhau. Một số trong số chúng, được gọi là đoạn, có vỏ mỏng và dễ thấm nước. Thông qua chúng, nước từ lớp vỏ sơ cấp đi vào bó dẫn hướng tâm của rễ. Các tế bào khác có độ dày xenlulo đặc biệt của các bức tường tiếp tuyến xuyên tâm và bên trong. Những nubs này được ngâm tẩm với suberin được gọi là đai Caspari. Chúng không thấm nước. Do đó, nước chỉ đi vào ống trụ trung tâm qua các ô dẫn. Và vì bề mặt hấp thụ của rễ vượt quá đáng kể tổng diện tích tiết diện của các tế bào đi qua của nội bì, khi đó áp suất rễ phát sinh, đây là một trong những cơ chế dẫn nước chảy vào thân, lá và các cơ quan sinh sản. .
Nội bì cũng là một phần của vỏ thân non. Ở một số thực vật hạt kín thân thảo, giống như rễ, có thể có các đai Caspari. Ngoài ra, ở thân non, nội bì có thể được biểu hiện bằng một vỏ bọc chứa nhiều tinh bột. Nhờ đó, nội bì có thể điều chỉnh sự vận chuyển nước trong cây và dự trữ chất dinh dưỡng.
6.6. Khái niệm về tấm bia và sự phát triển của nó
Người ta chú ý nhiều đến sự xuất hiện, phát triển của ontogeny và sự biến đổi cấu trúc tiến hóa của hệ thống dẫn, vì nó cung cấp sự liên kết giữa các cơ quan thực vật và sự tiến hóa của các đơn vị phân loại lớn gắn liền với nó.
Theo gợi ý của các nhà thực vật học người Pháp F. Van Thiegem và A. Dulio (1886), tập hợp các mô dẫn sơ cấp, cùng với các mô khác nằm giữa chúng và chu trình liền kề với vỏ cây, được gọi là tấm bia. Thành phần của tấm bia cũng có thể bao gồm một lõi và một khoang được hình thành ở vị trí của nó, chẳng hạn như bằng cỏ xanh. Khái niệm "tấm bia" tương ứng với khái niệm "hình trụ trung tâm". Tấm bia của rễ và thân có chức năng như nhau. Việc nghiên cứu tấm bia ở các đại diện của các bộ phận khác nhau của thực vật bậc cao đã dẫn đến sự hình thành lý thuyết về tấm bia.
Có hai loại bia chính: protostela và eustela. Cổ xưa nhất là protostela. Các mô dẫn của nó nằm ở giữa các cơ quan trục, với một xylem ở trung tâm được bao quanh bởi một lớp phloem liên tục. Không có lõi hoặc khoang trong thân.
Có một số loại protostela có liên quan về mặt tiến hóa: haplostela, actinostela và plectostela.
Loài nguyên thủy, nguyên thủy là haplostela. Xylem của cô ấy có hình dạng mặt cắt ngang tròn và được bao quanh bởi một lớp phloem liên tục. Xung quanh các mô dẫn điện trong một hoặc hai lớp là vòng tròn [K. Esau, 1969]. Haplostela được biết đến trong số các tê giác hóa thạch và được bảo tồn trong số một số loài psilotophytes (tmezipter).
Một loại protostela phát triển hơn là actinostela, trong đó xylem là mặt cắt ngang có dạng một ngôi sao nhiều tia. Nó được tìm thấy trong hóa thạch asteroxylon và một số động vật chân không nguyên thủy.
Sự tách rời xa hơn của xylem thành các khu vực riêng biệt, nằm hướng tâm hoặc song song với nhau, dẫn đến sự hình thành của một tấm đệm, đặc trưng của thân cây bạch huyết. Ở actinostela và plectostela, phloem vẫn bao quanh xylem từ mọi phía.
Trong quá trình tiến hóa, một siphonostel phát sinh từ protostela, một đặc điểm nổi bật của nó là cấu trúc hình ống của nó. Ở trung tâm của một tấm bia như vậy là một lõi hoặc một cái hốc. Trong phần dẫn điện của ống siphonostela, các khe lá xuất hiện, do đó xảy ra sự kết nối liên tục của lõi với vỏ. Tùy thuộc vào phương pháp sắp xếp lẫn nhau của xylem và phloem, siphonostela có thể là ectofloid và amphifloic. Trong trường hợp đầu tiên, phloem bao quanh xylem ở một mặt bên ngoài. Trong cách thứ hai, phloem bao quanh xylem từ hai phía, từ bên ngoài và từ bên trong.
Khi phân chia các siphonostela lưỡng tính thành một mạng lưới hoặc các hàng sợi dọc, một tấm bia bị chia cắt, hay còn gọi là dictyostela, đặc trưng của nhiều con giống dương xỉ, sẽ xuất hiện. Phần dẫn điện của nó được biểu diễn bằng nhiều bó dẫn điện đồng tâm.
Ở đuôi ngựa, một dấu hiệu khớp đã phát sinh từ siphonostela ectofloic, có cấu trúc liên kết. Nó được phân biệt bởi sự hiện diện của một khoang trung tâm lớn và các bó dẫn riêng biệt với các khoang protoxylem (ống tủy).
Ở thực vật có hoa, trên cơ sở một siphonostela ectofloid, một eustela, đặc trưng của cây hai lá mầm và một atactostela, điển hình của một lá mầm, đã được hình thành. Trong eustela, phần dẫn điện bao gồm các bó thế chấp riêng biệt với sự sắp xếp hình tròn. Ở trung tâm của tấm bia trong thân cây là lõi, được kết nối với vỏ cây với sự trợ giúp của các tia lõi. Trong atactostelle, các chùm dẫn có sự sắp xếp phân tán, giữa chúng là các tế bào nhu mô của hình trụ trung tâm. Sự sắp xếp này của các dầm che giấu cấu trúc hình ống của siphonostela.
Sự xuất hiện của nhiều biến thể khác nhau của siphonostela là một sự thích nghi quan trọng của thực vật bậc cao với sự gia tăng đường kính của các cơ quan trục - rễ và thân.
25. 8.1. Hệ bài tiết thực vật và ý nghĩa của nó
Đời sống thực vật là một tập hợp các phản ứng sinh hóa được xác định về mặt di truyền, tốc độ và cường độ của chúng phần lớn bị thay đổi bởi các điều kiện của môi trường phát triển. Trong các phản ứng này, nhiều loại sản phẩm phụ được hình thành không được thực vật sử dụng để xây dựng cơ thể hoặc để điều hòa sự trao đổi chất, năng lượng và thông tin với môi trường. Các sản phẩm như vậy có thể được loại bỏ khỏi cây theo nhiều cách khác nhau: trong quá trình chết đi và tách các cành và các phần của thân rễ, khi lá rụng và các lớp bên ngoài của lớp vỏ bong ra, do hoạt động của các cấu trúc chuyên biệt của bên ngoài và nội tiết. Cùng với nhau, những sự thích nghi này tạo thành hệ thống bài tiết của thực vật.
Không giống như động vật, hệ bài tiết ở thực vật không nhằm mục đích loại bỏ các hợp chất nitơ, có thể được tái sử dụng trong quá trình sống.
Hệ bài tiết của cây đa chức năng. Trong cấu trúc của nó được thực hiện: tổng hợp, tích lũy, dẫn truyền và giải phóng các sản phẩm trao đổi chất. Ví dụ, trong các tế bào tiết của ống dẫn nhựa ở lá cây lá kim, nhựa được hình thành, nhựa này được giải phóng qua ống dẫn nhựa. Trong mật hoa của hoa bằng lăng, mật hoa dịch ngọt được hình thành và tiết ra. Trái cây có múi tích tụ tinh dầu trong những chất chứa đặc biệt trong vỏ trái cây.
Sự hình thành và bài tiết các sản phẩm phụ của quá trình trao đổi chất có ý nghĩa thích nghi nhiều mặt:
Thu hút côn trùng thụ phấn. Trong hoa của táo, dưa chuột và các loài côn trùng thụ phấn chéo khác, mật hoa được hình thành, thu hút ong, và dịch tiết của hoa rafflesia thu hút ruồi;
Xua đuổi động vật ăn cỏ (thìa là, cây tầm ma, v.v.);
Bảo vệ chống lại vi khuẩn và nấm phá hủy gỗ (thông, vân sam, v.v.);
Giải phóng các hợp chất dễ bay hơi vào khí quyển, giúp làm sạch không khí khỏi vi khuẩn gây bệnh;
Sự tiêu hóa con mồi ngoại bào ở thực vật ăn côn trùng do giải phóng các enzym phân giải protein (sundew, aldrovanda, v.v.);
Sự khoáng hóa các tàn dư hữu cơ trong đất do rễ tiết ra các enzym đất đặc biệt;
Quy định chế độ nước bằng lỗ khí khổng - các lỗ thoát nước nằm dọc theo mép phiến lá (dâu, bắp cải, bà béo, v.v.);
Điều chỉnh sự bay hơi nước do giải phóng các hợp chất ete dễ bay hơi, làm giảm độ trong suốt và tính dẫn nhiệt của không khí gần bề mặt lá (cây lá kim);
Quy định chế độ muối của tế bào (marin, quinoa, v.v.);
Những thay đổi về tính chất hóa học và vật lý của đất, cũng như quy định về thành phần loài của hệ vi sinh đất dưới ảnh hưởng của chất tiết ra từ rễ;
Quy định sự tương tác của thực vật trong phytocenosis bằng các chất tiết ra từ rễ, thân và lá, được gọi là allelopathy (hành, tỏi, v.v.).
Các chất do thực vật tiết ra rất đa dạng. Bản chất của chúng phụ thuộc vào kiểu gen của cây.
Nhiều loài tiết ra nước (dâu tây, bắp cải), muối (pho mát, quinoa), monosaccharide và axit hữu cơ (bồ công anh, rau diếp xoăn), mật hoa (bồ đề, kiều mạch), axit amin và protein (dương, liễu), tinh dầu (bạc hà, hoa hồng. ), balsam (linh sam), nhựa (thông, vân sam), cao su (hevea, kok-sagyz), chất nhầy (tế bào nắp rễ, hạt sưng các loại cây khác nhau), dịch tiêu hóa (sundew, zhiryanka), chất lỏng độc (tầm ma, hogweed) và các hợp chất khác.
26.1.1. Phân cực
Sự phân cực là sự hiện diện của những khác biệt về sinh hóa, chức năng và cấu trúc ở các phần đối diện nhau về mặt đường kính của các cơ quan của một cơ thể thực vật toàn vẹn. Tính phân cực ảnh hưởng đến cường độ của các quá trình sinh hóa trong tế bào và hoạt động chức năng của các bào quan, quyết định thiết kế của các cấu trúc giải phẫu. Ở cấp độ sinh vật thực vật toàn vẹn, cực gắn liền với hướng sinh trưởng và phát triển trong điều kiện có tác dụng của lực hấp dẫn.
Hiện tượng phân cực được quan sát thấy ở các cấp độ tổ chức thực vật khác nhau. Ở cấp độ phân tử, nó thể hiện trong cấu trúc phân tử các chất hữu cơ, chủ yếu là axit nucleic và protein. Vì vậy, độ phân cực của sợi DNA được xác định bởi một trật tự đặc biệt của việc nối các nucleotide của nó. Sự phân cực của các polypeptit gắn liền với sự có mặt của nhóm amino –NH 2 và nhóm cacboxyl –COOH trong thành phần amino axit. Sự phân cực của các phân tử diệp lục là do sự hiện diện của một nhân porphyrin tetrapyrrole trong đó và dư lượng rượu - metanol và phytol.
Tế bào và các bào quan của chúng có thể có cực. Ví dụ, tại cực tái sinh của phức hợp Golgi, các thể đơn bào mới được hình thành, và tại cực tiết, các túi được hình thành, có liên quan đến việc bài tiết các sản phẩm trao đổi chất ra khỏi tế bào.
Sự phân cực của tế bào xảy ra trong quá trình hình thành chúng. Sự phân cực tế bào xảy ra do độ pH, điện tích và thế thẩm thấu, nồng độ O 2 và CO 2, các cation canxi, các chất hoạt động sinh lý và các nguyên tố dinh dưỡng khoáng. Nó cũng có thể phát sinh dưới tác động của áp suất cơ học, lực căng bề mặt và ảnh hưởng của các ô lân cận.
Phân cực tế bào là đối xứng và không đối xứng. Phân cực đối xứng là điều kiện tất yếu để phân chia nguyên phân và hình thành các tế bào con tương đương. Đặc biệt, các vi ống tế bào di chuyển đến mặt phẳng xích đạo và tham gia vào quá trình hình thành phragmoplast; nhiễm sắc thể phân chia và các crômatit của chúng phân kỳ về các cực đối diện của tế bào với sự trợ giúp của các sợi kéo của trục chất nhiễm sắc. Tại đây ribosome và ti thể bắt đầu tập trung. Các phức hợp Golgi di chuyển về phía trung tâm và được định hướng cụ thể trong không gian. Các cực bài tiết của chúng hướng về xích đạo của tế bào đang phân chia, điều này đảm bảo sự tham gia tích cực của organoid này vào việc hình thành lớp đệm giữa và màng tế bào sơ cấp.
Phân cực không đối xứng là đặc điểm của các tế bào chuyên biệt. Vì vậy, trong một quả trứng trưởng thành, nhân được dịch chuyển về cực hướng đến chalase, noãn và một không bào lớn nằm ở cực vi chất.
Sự phân cực không đối xứng càng rõ nét ở noãn được thụ tinh - hợp tử. Trong đó, nhân, được bao quanh bởi các ống của lưới nội chất, nằm ở mặt chalazal của tế bào. Ở đây, các phần tử của bộ xương được định vị dày đặc hơn, mật độ quang học của tế bào chất tăng lên, hàm lượng các enzym, phytohormon và các chất hoạt động sinh lý khác được ghi nhận tăng lên. Phần hoạt động nhiều hơn về mặt chức năng của nó trở thành đỉnh, và phần đối diện trở thành đáy.
Từ tế bào đỉnh, trong quá trình phát triển phôi của hạt, một cuống phôi có hình nón đang phát triển và các lá sơ khai được hình thành. Chất lơ lửng được hình thành từ tế bào đáy, và sau đó - rễ cột sống.
Do sự phân cực của các tế bào của phôi đang phát triển, hạt trưởng thành cũng phân cực. Ví dụ, ở hạt lúa mì, một mặt có nội nhũ và mặt khác là phôi, trong đó mô phân sinh đỉnh của thân và đỉnh của rễ nằm ở hai cực đối nhau.
Ngược lại với tế bào, sự phân cực của các cơ quan thực vật phổ biến hơn. Nó thể hiện trong cấu trúc của hệ thống chồi và rễ, cũng như các bộ phận cấu thành của chúng. Ví dụ, các phần đáy và đỉnh của chồi khác nhau về hình thái, giải phẫu, mô học, sinh hóa và chức năng. Ở nhiều loài thực vật có hoa, có một chồi đỉnh ở đỉnh của chồi, trong đó trung tâm phát sinh hình thái quan trọng nhất của thực vật - hình nón sinh trưởng. Nhờ hoạt động của nó, các lá thô sơ, chồi nách, nút và lóng được hình thành. Sự biệt hóa các tế bào của mô phân sinh sơ cấp dẫn đến sự xuất hiện của các mô nguyên sinh sơ cấp, các yếu tố mô học của vỏ sơ cấp và trụ trung tâm.
Đầu chồi là trung tâm thu hút mạnh mẽ. Đây là nơi dẫn dòng chảy chính của nước và các nguyên tố hòa tan dinh dưỡng khoáng và các chất hữu cơ được tổng hợp ở rễ. Sự cung cấp chủ yếu của cytokinin cho đỉnh thận dẫn đến sự chiếm ưu thế ở đỉnh. Tế bào của đỉnh chồi ngọn đang phân chia tích cực, đảm bảo sự phát triển chiều dài của thân và hình thành các lá mới và chồi nách trên trục chính. Trong trường hợp này, có một sự ức chế tương quan đối với sự phát triển của chồi nách.
Một ví dụ điển hình về sự phân cực hình thái chồi là cấu trúc của thân lúa mì. Khi bạn di chuyển từ dưới lên trên, từ phần gốc của chồi đến ngọn, các lóng trở nên dài hơn, độ dày của chúng ở phần giữa đầu tiên tăng lên và sau đó giảm dần. Ở các lá lớp trên, tỉ số giữa chiều rộng của phiến lá và chiều dài của nó lớn hơn nhiều so với các lá ở lớp dưới. Vị trí của lá trong không gian cũng thay đổi. Ở lúa mì, các lá của tầng dưới rủ xuống, các lá của tầng giữa nằm gần như plagiotropical, tức là. song song với bề mặt trái đất, và các lá cờ (trên) có xu hướng nằm ở vị trí trực hướng - gần như thẳng đứng.
Độ phân cực của cú đánh được xác định rõ ràng ở cấp độ giải phẫu. Ở các loài lúa mì, các lóng dương, so với các lóng có vị trí thấp hơn, được phân biệt bởi đường kính nhỏ hơn, số lượng bó dẫn ít hơn và nhu mô đồng hóa phát triển tốt hơn.
Ở thực vật hai lá mầm, tính phân cực giải phẫu của chồi được tăng cường nhờ sự xuất hiện của các mô giáo dục bên thứ cấp - cambium và phellogen. Phellogen tạo ra một mô bao phủ thứ cấp của phelleme, được ngâm tẩm với suberin, biến thành nút chai. Ở các loài cây, hoạt động của phellogen dẫn đến sự hình thành phức hợp bao phủ bậc ba - lớp vỏ. Cambium cung cấp một sự chuyển đổi sang cấu trúc giải phẫu thứ cấp của thân dưới mức đỉnh thận.
Rễ, cũng như các cơ quan khác của thực vật, cũng được đặc trưng bởi sự phân cực về cấu trúc và chức năng. Do địa chất dương nên phần gốc của rễ nằm ở bề mặt đất. Nó liên quan trực tiếp đến cổ rễ - nơi mà rễ đi vào thân. Phần đỉnh của rễ thường được chôn trong đất. Các quá trình tăng trưởng và phát triển được kết nối với nhau diễn ra trong đó. Sự phân chia tế bào của mô phân sinh đỉnh tạo ra sự phát triển tuyến tính của rễ. Và kết quả của sự biệt hóa tế bào, các cấu trúc mới về chất lượng được hình thành. Lông rễ xuất hiện trên các tế bào của biểu bì. Các phần tử của vỏ não sơ cấp được hình thành từ các tế bào của chu vi, và vòng tròn và các phần tử dẫn điện của hình trụ trung tâm được hình thành từ màng phổi. Thiết kế vùng rễ non này đảm bảo khả năng hấp thụ tích cực nước và khoáng chất, cũng như cung cấp cho vùng rễ cao hơn. Các chức năng của phần cơ bản của rễ có phần khác nhau. Vì vậy, ở cây hai lá mầm lâu năm, phần đáy của rễ thực hiện chức năng vận chuyển, nâng đỡ và lưu trữ. Cấu trúc giải phẫu tương ứng với việc thực hiện các chức năng này. Các mô dẫn được phát triển tốt hơn ở đây, vỏ não được hình thành do hoạt động của cambium, mô liên kết được đại diện bởi một nút bần. Sự phân cực của cấu trúc rễ đảm bảo tính đa dạng trong các chức năng của nó.
Tính phân cực cũng là đặc điểm của cơ quan sinh sản của thực vật. Vì vậy, hoa, là một chồi rút ngắn đã được sửa đổi, vẫn giữ các dấu hiệu của sự phân cực của chồi. Các bộ phận của hoa nằm trên đó theo một trình tự đều đặn: đài hoa, tràng hoa, androecium và gynoecium. Sự sắp xếp này thúc đẩy quá trình thu phấn hoa tốt hơn nhờ đầu nhụy, và cũng bảo vệ các bộ phận sinh dưỡng của hoa bởi các bộ phận sinh dưỡng. Sự phân cực của chùm hoa rất rõ ràng. Trong các cụm hoa không xác định, các hoa của phần gốc được đặt trước. Chúng phát triển với kích thước lớn và tạo ra quả với hạt phát triển tốt. Đặc biệt, các hạt lớn hơn trong hướng dương được hình thành ở phần ngoại vi của giỏ, và trong một tai phức tạp của lúa mì hạt giống tốt nhấtđược hình thành trong những bông hoa đầu tiên của bông hoa. Hoa của phần ngọn của cụm hoa được hình thành muộn hơn. Quả và hạt thu được từ chúng nhỏ hơn và chất lượng gieo hạt thấp hơn.
Vì vậy, tính phân cực là một đặc điểm cấu tạo, chức năng và sinh hóa quan trọng của thực vật có giá trị thích nghi và phải được tính đến trong thực hành nông học.
1.2. Đối diện
Thế giới xung quanh chúng ta được đặc trưng bởi tính toàn vẹn và trật tự hài hòa. Trong không gian bên ngoài gần nhất, vị trí, khối lượng, hình dạng và quỹ đạo chuyển động của các vật thể là hài hòa. Hệ mặt trời... Trên Trái đất, sự thay đổi theo mùa và hàng ngày của các thông số vật lý quan trọng nhất của điều kiện tồn tại của các sinh vật là hài hòa, cũng được đặc trưng bởi sự phối hợp hài hòa của cấu trúc và chức năng. Các nhà hiền triết của trường phái Pythogor tin rằng sự hài hòa là "một cách phối hợp nhiều bộ phận với nhau, với sự giúp đỡ mà chúng hợp nhất thành một tổng thể." Đối xứng là sự phản ánh của sự hài hòa trong tự nhiên. Theo Yu.A. Đối xứng Urmantsev là một phạm trù biểu thị sự bảo toàn các tính năng của đối tượng liên quan đến sự thay đổi của chúng. Theo thuật ngữ thực dụng, đối xứng là tính đồng nhất của cấu trúc và sự sắp xếp lẫn nhau của cùng một loại các bộ phận cấu thành của một tổng thể duy nhất. Tính đối xứng vốn có trong cả khoáng chất và động vật hoang dã. Tuy nhiên, các dạng đối xứng và mức độ biểu hiện của chúng trong các đối tượng khác nhau có sự khác biệt đáng kể.
Một đặc điểm thiết yếu của tính đối xứng của các đối tượng có nguồn gốc khác nhau là đặc tính quan hệ của nó. So sánh các cấu trúc được thực hiện trên một số điểm đặc trưng. Các điểm của hình cho cùng một hình khi nhìn hình từ các phía khác nhau được gọi là bằng nhau. Đó có thể là các điểm cách đều nhau trên một đường thẳng, giao điểm của các cạnh của tam giác cân và hình vuông, các mặt của hình đa diện, các điểm trên đường tròn hoặc trên bề mặt của một quả bóng, v.v.
Nếu không có điểm nào bằng nhau trong hình đối với điểm X nào đó thì điểm đó được gọi là đặc biệt. Các hình có một điểm đặc biệt và một số điểm bằng nhau được gọi là hình hoa thị. Các hình không chứa các điểm bằng nhau được coi là không đối xứng.
Các trục và mặt phẳng đối xứng đi qua các điểm kỳ dị. Trong một tam giác cân, ba có thể được vẽ trong một hình vuông - bốn, trong một ngũ giác đều - năm mặt phẳng đối xứng. Theo đó, hai chùm đối xứng được hình thành trên một đường thẳng, đối xứng ba chùm là đặc trưng của tam giác, đối xứng bốn chùm là đặc trưng của hình vuông và đối xứng nhiều chùm là đặc trưng của đường tròn.
Các chuyển động của một hình, do mỗi điểm được thay thế bằng một điểm bằng nhau, và mỗi điểm đơn lẻ vẫn giữ nguyên vị trí, được gọi là các phép biến đổi đối xứng và các hình cho phép phép biến đổi đối xứng được coi là đối xứng.
Các dạng biến đổi đối xứng phổ biến nhất là:
1. THAM KHẢO - chuyển động của mỗi điểm nằm cách mặt phẳng cố định một khoảng nào đó, trên đường thẳng vuông góc với mặt phẳng này, cùng khoảng cách ở mặt kia của nó (ví dụ, đối xứng qua gương của hoa đậu biếc);
2. P khoảng trong khoảng t - chuyển động của tất cả các điểm theo một góc xác định quanh một trục cố định (ví dụ, đối xứng đa chùm của một bông hoa anh đào hoạt hóa);
3. Các lông tơ ở chân, ví dụ, vị trí của các lông tơ trên chồi.
Các dạng biến đổi đối xứng không đồng nhất và không giảm xuống nhau. Do đó, trong trường hợp phản xạ, máy bay vẫn đứng yên; khi quay - đường thẳng (trục); và với phép tịnh tiến song song, không có điểm nào được giữ nguyên.
Hình đối xứng có một số điểm bằng nhau có thể có một số điểm đặc biệt khác nhau. Vì vậy, hoa thị có một điểm đặc biệt. Hình thức chính của phép biến đổi đối xứng cho nó là phép quay. Đối với các hình không có điểm kỳ dị, một phép dời hình song song, hoặc sự dịch chuyển, là đặc trưng. Những số liệu như vậy được quy ước là vô tận.
2.2.1. Đặc điểm về sự biểu hiện của tính đối xứng ở thực vật. Tính đối xứng của thực vật khác với đối xứng của tinh thể ở một số đặc điểm.
1) Tính đối xứng của thực vật không chỉ được xác định bởi sự đối xứng của các phân tử tạo thành tế bào của chúng, mà còn bởi sự đối xứng của các điều kiện môi trường trong đó diễn ra quá trình phát triển của thực vật.
Sự đồng nhất về thành phần của lớp rễ của đất, sự đồng đều của sự phân bố nước và các nguyên tố dinh dưỡng khoáng trong đất, và sự xa nhau bằng nhau của các cây mọc gần đó có thể được coi là những trường hợp đối xứng cụ thể của môi trường sống. Những trường hợp này phải được tính đến khi trồng cây trồng.
2) Thực vật, giống như các sinh vật sống khác, không có sự đồng nhất tuyệt đối về các yếu tố của các bộ phận cấu thành của chúng. Điều này chủ yếu được xác định bởi thực tế là các điều kiện để hình thành các bộ phận này không hoàn toàn giống nhau. Một mặt, điều này là do sự khác biệt về thời gian phân tách hình nón sinh trưởng của các metameres khác nhau, bao gồm các lá, chồi, nút và lóng thô sơ. Điều tương tự cũng áp dụng cho các cơ quan sinh sản. Như vậy, những bông hoa hình ống ở rìa trong lẵng hoa hướng dương luôn lớn hơn những bông hoa ở phần trung tâm của chùm hoa. Mặt khác, trong mùa sinh trưởng, cả những thay đổi thường xuyên và ngẫu nhiên của các điều kiện phát triển đều được quan sát thấy. Ví dụ, nhiệt độ của không khí và đất, độ chiếu sáng và thành phần quang phổ của ánh sáng thay đổi một cách thống kê thường xuyên trong mùa sinh trưởng. Những thay đổi cục bộ trong việc cung cấp chất dinh dưỡng trong đất, ảnh hưởng của dịch bệnh và sâu bệnh có thể là ngẫu nhiên.
Sự thiếu vắng sự đồng nhất tuyệt đối của các bộ phận cấu thành có tầm quan trọng lớn trong đời sống của thực vật. Tính không đồng nhất kết quả là một trong những cơ chế cho độ tin cậy của quá trình phát sinh.
3) Thực vật được đặc trưng bởi sự đối xứng về cơ bản, không gian và thời gian.
A. Phép đo cơ bản bao gồm sự lặp lại chính xác của hình dạng và các thông số tuyến tính của cấu trúc, cũng như sự thay đổi chính xác của các thông số này. Nó là đặc trưng của tinh thể các chất vô cơ, phân tử hợp chất hữu cơ, bào quan tế bào, cấu trúc giải phẫu và cơ quan thực vật. Cấp độ cao Các phân tử DNA khác nhau về tính đối xứng đáng kể. Khi độ ẩm của chế phẩm gần bằng sinh lý, phân tử DNA ở dạng B và được đặc trưng bởi sự lặp lại rõ ràng các thông số của nó. các yếu tố cấu thành... Mỗi vòng xoắn của phân tử ADN chứa 10 nuclêôtit; hình chiếu của cuộn dây lên trục của phân tử là 34,6 Å (1 Å = 1 · 10 –10 m); khoảng cách chiếu giữa các nucleotide liền kề là 3,4 Å và khoảng cách tuyến tính là 7 Å; đường kính của phân tử khi định hướng dọc theo các nguyên tử photpho gần bằng 20 Å; đường kính của rãnh lớn là khoảng 17 Å và đường kính của rãnh nhỏ là 11 Å.
Tính đối xứng đáng kể của thực vật không chỉ được đặc trưng bởi sự lặp lại chính xác của các thông số của cấu trúc, mà còn bởi sự thay đổi thường xuyên của chúng. Ví dụ, ở cây vân sam, sự giảm đường kính thân trên 1 m chiều dài, khi di chuyển từ phần gốc của nó lên phần ngọn, là tương đối không đổi. Ở cây dong riềng tím (Canna violacea), hoa theo truyền thống được coi là không đối xứng. Các lá đài, cánh hoa và nhị của nó có kích thước khác nhau. Tuy nhiên, sự thay đổi kích thước tuyến tính của các thành viên này của hoa vẫn tương đối không đổi, đó là một dấu hiệu của sự đối xứng. Chỉ ở đây, thay vì gương một, các dạng đối xứng khác phát triển.
B. Chiều không gian bao gồm sự sắp xếp không gian lặp lại thường xuyên của các bộ phận cấu thành giống nhau ở thực vật. Đối xứng không gian phổ biến rộng rãi trong thế giới thực vật. Nó là điển hình cho vị trí của chồi trên chồi, hoa trong chùm hoa, hoa viên trên bao, phấn hoa trong bao phấn, vảy trong nón hạt trần và nhiều trường hợp khác. Một trường hợp đặc biệt của đối xứng không gian là đối xứng dọc, xuyên tâm và hỗn hợp của thân cây.
Đối xứng dọc phát sinh trong quá trình truyền song song, tức là sự lặp lại không gian của metameres trong cấu trúc chồi. Nó được xác định bởi sự giống nhau về mặt cấu tạo của các bộ phận cấu thành của chồi - metameres, cũng như sự tương ứng của chiều dài các lóng với quy tắc của "phần vàng".
Một trường hợp phức tạp hơn về đối xứng không gian là sự sắp xếp của các lá trên chồi, có thể mọc thành chùm, đối diện và xen kẽ (xoắn ốc).
Đối xứng xuyên tâm phát sinh khi trục cấu trúc kết hợp với các mặt phẳng đối xứng đi qua nó. Đối xứng xuyên tâm phổ biến trong tự nhiên. Đặc biệt, là đặc trưng của nhiều loài tảo cát, là mặt cắt ngang thân của cây sinh bào tử bậc cao, cây hạt trần và cây hạt kín.
Đối xứng xuyên tâm của chồi là đa yếu tố. Nó đặc trưng cho vị trí của các bộ phận cấu thành và có thể được biểu thị bằng các chỉ số chiều khác nhau: khoảng cách của các bó dẫn và các cấu trúc giải phẫu khác từ điểm kỳ dị mà một hoặc nhiều mặt phẳng đối xứng đi qua, nhịp điệu của sự luân phiên của các cấu trúc giải phẫu, góc phân kỳ, cho thấy sự dịch chuyển của trục của một cấu trúc so với trục của cấu trúc kia. Ví dụ, trong một bông hoa anh đào, năm cánh hoa tràng hoa được sắp xếp đối xứng nhau về mặt không gian. Nơi gắn của mỗi bông hoa vào lọ ở cùng một khoảng cách từ tâm của bông hoa, và góc phân kỳ sẽ bằng 72º (360º: 5 = 72º). Đối với một bông hoa tulip, góc phân kỳ của mỗi trong số sáu cánh hoa là 60º (360º: 6 = 60º).
C. Chiều thời gian của thực vật thể hiện ở sự lặp lại nhịp nhàng theo thời gian của các quá trình phát sinh hình thái và các chức năng sinh lý khác. Ví dụ, sự phân tách sự phát triển của lá nguyên thủy theo hình nón được thực hiện trong những khoảng thời gian bằng nhau hoặc ít hơn, được gọi là plastochron. Những thay đổi theo mùa trong các quá trình hoạt động sống của cây đa thân lâu năm được lặp lại rất nhịp nhàng. Đối xứng thời gian phản ánh sự thích nghi của thực vật với những thay đổi hàng ngày và theo mùa của điều kiện môi trường.
4) Tính đối xứng của cây phát triển linh động trong quá trình hình thành và đạt đến biểu hiện tối đa trong quá trình sinh sản hữu tính. Ở thực vật, một ví dụ về sự hình thành đối xứng xuyên tâm của chồi là biểu hiện về mặt này. Ban đầu, các tế bào của nón sinh trưởng ít nhiều đồng nhất, không biệt hóa. Do đó, rất khó về mặt hình thái học để phân biệt một tế bào hoặc một nhóm tế bào qua đó có thể vẽ một mặt phẳng đối xứng.
Sau đó, các tế bào của áo dài tạo thành lớp biểu bì, từ đó lớp biểu bì được hình thành. Đồng hóa và lưu trữ các mô, cũng như các mô cơ giới sơ cấp và lõi, phát triển từ mô phân sinh chính. Ở vùng ngoại vi, các sợi tế bào procambium hẹp và dài được hình thành, từ đó các mô dẫn truyền sẽ phát triển. Với sự hình thành liên tục của procambium, các lớp phloem và xylem liên tục được hình thành từ nó. Nếu procambium được đặt ở dạng sợi, thì các bó dẫn riêng biệt được hình thành từ nó. Ở các cây thân gỗ, hoạt động chức năng lâu năm, thay đổi theo mùa của cambium sẽ dẫn đến sự hình thành của vỏ thứ cấp và các vòng gỗ hàng năm, điều này sẽ tăng cường tính đối xứng hướng tâm của thân cây.
5) Tính đối xứng của thực vật thay đổi trong quá trình tiến hóa. Các phép biến đổi tiến hóa của phép đối xứng có tầm quan trọng lớn trong sự phát triển của thế giới hữu cơ. Do đó, sự xuất hiện của đối xứng hai bên là một sự thích nghi lớn về mặt sinh lý học (tính thơm) đã nâng cao đáng kể mức độ tổ chức của động vật. Trong giới thực vật, sự thay đổi số lượng các kiểu đối xứng cơ bản và các dẫn xuất của chúng có liên quan đến sự xuất hiện của đa bào và sự xuất hiện của thực vật trên cạn. Ở tảo đơn bào, đặc biệt là tảo cát, hiện tượng đối xứng xuyên tâm là phổ biến. Ở các sinh vật đa bào, nhiều dạng chuyển giao song song khác nhau xuất hiện, cũng như các dạng đối xứng hỗn hợp. Ví dụ, ở tảo charoh, tính đối xứng xuyên tâm của mặt cắt thallus được kết hợp với sự hiện diện của trục chuyển metameric.
Sự tiến hóa của hệ thống dẫn điện có tầm quan trọng lớn trong việc hình thành tính đối xứng của thực vật bậc cao. Protostele thân đối xứng xuyên tâm của các dạng nguyên thủy được thay thế bằng một phức hợp các bó thế hệ đối xứng hai bên (đơn đối xứng) tạo thành eustela và đảm bảo tính đối xứng xuyên tâm của thân ở thực vật hạt kín.
Xu hướng chung trong sự tiến hóa của tính đối xứng trong các cơ thể sống, bao gồm cả thực vật, là sự giảm mức độ đối xứng. Điều này là do sự giảm số lượng các loại đối xứng cơ bản và các dẫn xuất của chúng. Vì vậy, các loài thực vật có hoa sớm hơn về mặt tiến hóa (họ Magnoliaceae, họ Buttercup, v.v.) được đặc trưng bởi các bộ phận đa thức, tự do, nằm ở vị trí xoắn ốc của hoa. Trong trường hợp này, hoa hóa ra có khả năng hoạt hóa, tức là không đối xứng. Công thức của những bông hoa như vậy có xu hướng biểu thức: Å Ca ¥ Co ¥ A ¥ G ¥ ... Trong quá trình tiến hóa, người ta quan sát thấy sự giảm số lượng hoa thành viên cũng như sự bồi tụ của chúng, điều này luôn dẫn đến việc giảm số lượng mặt phẳng đối xứng.
Ví dụ, trong các gia đình trẻ tiến hóa, ở cừu non hoặc cỏ xanh, hoa trở nên đa hình (đơn đối xứng). Sự bất đối xứng của hoa rõ ràng có một ý nghĩa thích nghi gắn liền với việc cải tiến các phương pháp thụ phấn. Điều này thường được tạo điều kiện thuận lợi bởi sự tiến hóa đồng thời của hoa và các loài thụ phấn - côn trùng và chim.
Sự đặc biệt của sự đối xứng của hoa trong chùm hoa có tầm quan trọng lớn. Vì vậy, trong một giỏ hướng dương, những bông hoa hợp tử biên vô sinh có một cái lưỡi giả lớn màu vàng, được tạo thành bởi ba cánh hoa hợp nhất. Các hoa màu mỡ của phần trung tâm của cụm hoa là đơn tính hoạt hóa, chúng được hình thành bởi năm cánh hoa nhỏ có kích thước bằng nhau, dồn lại thành một ống. Một ví dụ khác là các đại diện của họ Celery. Ở họ, trong một ô phức tạp, các hoa rìa có khả năng sinh hợp tử yếu, trong khi các hoa khác vẫn có khả năng hoạt hóa điển hình.
Sự tiến hóa của cấu trúc và tính đối xứng của thực vật không đơn giản. Sự bất đối xứng, chiếm ưu thế về vai trò của nó, được thay thế và bổ sung ở những giai đoạn tiến hóa nhất định bằng sự đối xứng.
Vì vậy, tính đối xứng của thực vật và các bộ phận cấu thành của chúng là rất nhiều mặt. Nó gắn liền với sự đối xứng của các yếu tố cấu thành nó ở cấp độ phân tử, tế bào, mô học-giải phẫu và hình thái học. Sự đối xứng phát triển linh hoạt trong quá trình phát sinh và phát sinh loài và tạo ra mối liên hệ giữa thực vật và môi trường.
1.3. Chủ nghĩa siêu phàm
Sự thích nghi về mặt sinh lý học quan trọng của thực vật là metamerism, là sự hiện diện của các cấu trúc hữu hạn cơ bản lặp lại, hay metameres, trong hệ thống của toàn bộ sinh vật. Cấu trúc số liệu cung cấp sự lặp lại nhiều lần của các phần cấu thành của cảnh quay và do đó, là một trong những cơ chế tạo nên độ tin cậy của quá trình hình thành. Cấu trúc metameric là đặc trưng của các nhóm thực vật phân loại khác nhau. Nó được biết đến trong tảo rừng ngập mặn, cỏ đuôi ngựa và các thực vật sinh bào tử bậc cao khác, thực vật hạt trần, thực vật hạt kín trên cạn và dưới nước. Các bộ phận khác nhau về mặt chức năng của thực vật là bộ phận sinh dưỡng - sinh dưỡng và sinh dưỡng. Với sự tham gia của các cơ quan được sắp xếp theo hệ metan trong thực vật, một hệ thống metame được hình thành. Sự phân nhánh là một trường hợp đặc biệt của sự hình thành các hệ mét.
Vùng sinh dưỡng của chồi cây hạt kín bao gồm một lá, một nút, một lóng và một chồi nách bên, nằm ở gốc của lóng ở phía đối diện với nơi bám của lá. Chồi này được bao phủ bởi một tấm của metamer trước đó. Các metameres của vùng sinh sản rất đa dạng. Ví dụ, ở lúa mì, metame của một cành hợp chất bao gồm một đoạn của bộ xương và một bộ xương gắn liền với nó. Đôi khi thực vật có vùng chồi chuyển tiếp. Ở một số loài và giống lúa mì, nó có thể được biểu thị bằng vảy của các bông nhỏ kém phát triển.
Độ lớn của thực vật là một biểu hiện hình thái về tính đặc trưng của quá trình sinh trưởng và phát sinh hình thái của chúng, diễn ra một cách nhịp nhàng, dưới dạng các chu kỳ phụ lặp đi lặp lại khu trú ở các ổ của mô phân sinh. Nhịp điệu của quá trình hình thành metame có liên quan chặt chẽ với tính chu kỳ của các quá trình sinh trưởng đặc trưng của thực vật. Sự hình thành các metameres là quá trình phát sinh hình thái cơ bản trong quá trình phát triển của thực vật. Nó tạo cơ sở cho sự phức tạp của tổ chức trong ontogeny và phản ánh quá trình trùng hợp, là một trong những cơ chế tiến hóa của thực vật bậc cao.
Sự hình thành và phát triển của cây mầm chủ yếu do chức năng của mô phân sinh ngọn và mô phân sinh giữa các lớp.
Trong chồi sinh dưỡng, cũng như trong phôi của hạt nảy mầm, do kết quả của quá trình phân bào, sự gia tăng thể tích nón sinh trưởng của cuống phôi xảy ra. Sự phân chia tích cực sau đó của các tế bào ở vùng ngoại vi của nón sinh trưởng dẫn đến sự hình thành nguyên sinh lá - nguyên sinh lá và đĩa chèn. Lúc này, hoạt động mô phân sinh của tua-bin và mô phân sinh vùng trung tâm giảm nhẹ, nhưng các tế bào của đĩa chèn đang phân chia tích cực. Phần trên của đĩa là nơi bám của lá nguyên sinh, và khi nó phát triển dày lên, một nút thắt được hình thành từ nó. Một lóng phát triển từ phần dưới của đĩa chèn. Ở đây, phần củ thô sơ của chồi nách bên được hình thành ở phía đối diện với gân giữa của lá nguyên sinh. Kết hợp với nhau, chồi lá, đĩa chèn và chồi chồi là một metamere phôi thai.
Khi quá trình hình thành phôi thai hoàn thành, hoạt động phân chia tế bào ở vùng áo và vùng mô phân sinh trung tâm lại tăng lên. Thể tích của phần nhẵn của nón sinh trưởng tăng trở lại, đạt cực đại trước khi hình thành lá nguyên thủy tiếp theo. Do đó, một cái mới bắt đầu hình thành trên phần đỉnh của metamere thô sơ trước đó. Quá trình này được xác định về mặt di truyền và lặp lại nhịp nhàng nhiều lần. Trong trường hợp này, metamer được hình thành đầu tiên sẽ nằm ở phần cơ bản của chồi và về mặt di truyền là phần trẻ nhất - ở phần đỉnh. Tích lũy số lượng nguyên sinh thiên thạch ở T.I. Serebryakova gọi đó là sự chín muồi. Bà đã đặt ra thuật ngữ "khả năng của thận" để chỉ số lượng metameres tối đa có thể được gửi vào thận.
Sự sinh trưởng và phát triển của các lóng với tư cách là bộ phận cấu thành của ngọn phần lớn là do hoạt động của mô phân sinh giữa các đốt. Sự phân chia của các tế bào của mô phân sinh này và sự kéo dài của các dẫn xuất của chúng dẫn đến sự kéo dài của các lóng.
Metameres có một số tính năng đặc trưng cho phép chúng đảm bảo tính toàn vẹn về cấu trúc và chức năng của cơ thể thực vật.
1) Sự phân cực của metamers. Mỗi metamere có một phần cơ bản và phần đỉnh. Thuật ngữ "phần đỉnh" gợi ý rằng phần trên của ngọn hoặc chứa mô phân sinh đỉnh hoặc hướng về đỉnh.
Phần đáy và phần đỉnh khác nhau về các đặc điểm hình thái, mô học - giải phẫu và sinh lý - sinh hóa. Ví dụ, ở lúa mì, khi di chuyển từ phần gốc đến phần đỉnh của lóng, độ dày của thân, đường kính của ống tủy và độ dày của vòng ngoại vi của bệnh xơ cứng đầu tiên tăng lên và sau đó giảm dần; số lượng tế bào trong các dây của nhu mô đồng hóa tăng lên đáng kể; đường kính xuyên tâm của các bó dẫn giảm, cũng như số lượng mạch trong xylem.
2) Tính đối xứng của metameres. Sự đối xứng của các cơ quan thực vật được đảm bảo bởi sự đối xứng của các metamers hình thành chúng, phát sinh do một trình tự phân chia tế bào cụ thể của nón sinh trưởng.
3) Sự hình thành dị thường của metamers. Các metameres ở các thời điểm khác nhau, được cách ly xen kẽ bởi một hình nón sinh trưởng. Do đó, các metamere đầu tiên của phần cơ bản của chồi già hơn về mặt di truyền và các metameres cuối cùng của phần ngọn trẻ hơn. Về mặt hình thái, chúng khác nhau về các thông số tấm, chiều dài và độ dày của các lóng. Ví dụ, trong ngũ cốc, các lá của tầng trên rộng hơn các lá của tầng dưới, và các lóng dưới cành dài hơn và mỏng hơn các lóng dưới.
4) Sự biến đổi của các tính trạng metamer. Các siêu sao trẻ hơn về mặt di truyền có biên độ biến thiên của các ký tự thấp hơn. Do đó, các đặc điểm giải phẫu của lóng dương có thể được sử dụng với độ chính xác cao hơn để xác định các giống cây ngũ cốc và biên soạn các chương trình nhân giống.
5) Tính tối ưu của thiết kế metamers. Thiết kế sinh học tối ưu là thiết kế yêu cầu một lượng chất hữu cơ tối thiểu để xây dựng và duy trì. Vì cấu trúc của thực vật gắn bó chặt chẽ với chức năng của chúng, tiêu chí của tính tối ưu có thể là khả năng thích ứng của thực vật với các điều kiện phát triển và tỷ lệ giữa năng suất hạt của chúng với khối lượng cơ quan sinh dưỡng. Tính tối ưu của thiết kế một nhà máy tích hợp được đảm bảo bởi thiết kế tối ưu của các metamers cấu thành của nó.
6) Toàn bộ cây là một hệ thống cao phân tử do một tập hợp các metan tạo thành. Tính đa bội thể hiện ở tất cả các cấp độ tổ chức của cơ thể thực vật. Vì vậy, trong cấu trúc của ADN có nhiều đoạn gen lặp lại; Karyotype của nhiều loài thực vật được biểu hiện bằng bộ nhiễm sắc thể lưỡng bội hoặc đa bội; v một số lượng lớn tế bào chứa plastids, ti thể, ribosome và các bào quan khác. Cây trưởng thành là tập hợp cấp dưới gồm chồi, rễ và cơ quan sinh sản có cấu tạo kiểu metan. Hệ thống đo lường đảm bảo năng suất cao của cây trồng và tăng đáng kể độ tin cậy của quá trình phát sinh của chúng.
27.4.1. Cấu trúc giải phẫu cơ bản của rễ
Các đặc điểm của cấu tạo sơ cấp được biểu hiện rõ ràng ở các mặt cắt dọc và ngang của ngọn rễ.
Trên mặt cắt dọc của ngọn rễ, có thể phân biệt 4 múi.
Vùng nắp rễ bao phủ các mô phân sinh ngọn. Nó được tạo thành từ các tế bào sống. Lớp bề mặt của chúng liên tục bong ra và tạo thành đường đi mà rễ di chuyển. Các tế bào bong tróc cũng tạo ra chất nhầy, giúp đầu rễ di chuyển trong đất dễ dàng hơn. Các tế bào của phần trung tâm của nắp, hay còn gọi là cột sống, chứa các hạt tinh bột, góp phần vào sự phát triển địa dưỡng của rễ. Các tế bào của nắp liên tục được đổi mới do sự phân chia tế bào của một mô giáo dục đặc biệt - caliptrogen, là đặc điểm của monocots.
I - vùng của nắp rễ; II - vùng tăng trưởng; III - vùng của các lông hút ở rễ; IV - vùng dẫn truyền. 1 - biểu bì, 2 - chu kỳ, 3 - nội bì, 4 - vỏ sơ cấp, 5 - ngoại bì, 6 - trụ trung tâm, 7 - lông hút; 8 - sự hình thành rễ bên.
Vùng tăng trưởng bao gồm hai vùng con. Trong phân bào của sự phân chia, sự phát triển của rễ được thực hiện do quá trình phân bào giảm nhiễm tích cực. Ví dụ, ở lúa mì, tỷ lệ tế bào phân chia (chỉ số phân bào) là 100-200 ppm. Phân vùng phân chia là một vật liệu có giá trị cho các nghiên cứu di truyền tế bào. Ở đây thuận tiện cho việc nghiên cứu số lượng, cấu trúc vĩ mô và vi mô của nhiễm sắc thể. Trong tiểu vùng kéo dài, hoạt động mô phân sinh của tế bào giảm, nhưng do sự cân bằng nhất định của phytohormon, chủ yếu là auxin và cytokinin, sự phát triển của rễ xảy ra do sự kéo dài theo trục của tế bào non.
Vùng hút có thể được gọi một cách chính xác là vùng của lông rễ, cũng như vùng phân hóa, vì epiblema, vỏ não sơ cấp và trụ trung tâm được hình thành ở đây.
Epiblema là một mô liên kết đặc biệt, liên tục được đổi mới, bao gồm hai loại tế bào. Các lông hút có vách mỏng dài 1 - 3 mm phát triển từ các nguyên bào trichoblast, nhờ đó mà quá trình hấp thụ nước và các chất hòa tan trong nó được thực hiện. Các lông hút ở rễ có thời gian tồn tại ngắn. Chúng sống từ 2 đến 3 tuần và sau đó rụng rời. Nguyên bào atrichoblasts không hình thành lông rễ và thực hiện chức năng bổ sung.
Vỏ não sơ cấp điều chỉnh dòng chảy của nước vào các mô dẫn của hình trụ trung tâm. Trong quá trình hình thành trụ trung tâm, các mô protofloem là mô đầu tiên được hình thành, qua đó các chất hữu cơ cần thiết cho sự phát triển của nó đi vào ngọn rễ. Phía trên rễ, ở cấp độ của các lông rễ đầu tiên, các phần tử mạch của protoxylem xuất hiện. Các mô mạch nguyên thủy phát triển thành phloem sơ cấp và xylem sơ cấp của bó mạch hướng tâm.
Vùng dẫn truyền chiếm phần lớn nhất của rễ. Nó thực hiện nhiều chức năng: vận chuyển nước và các chất hữu cơ và khoáng hòa tan, tổng hợp các hợp chất hữu cơ, lưu trữ các chất dinh dưỡng, v.v. Vùng dẫn kết thúc bằng cổ rễ, tức là. nơi chuyển tiếp của rễ lên thân. Ở thực vật hạt kín hai lá mầm, ở phần đầu của đới này, có sự chuyển tiếp từ cấu trúc giải phẫu sơ cấp sang thứ cấp của rễ.
Trên mặt cắt ngang trong vùng lông hút của rễ, các đặc điểm cấu tạo và địa hình của cấu trúc giải phẫu cơ bản của rễ được bộc lộ (Hình 2). Trên bề mặt của rễ có một biểu bì với các lông hút của rễ, với sự trợ giúp của nước và các chất hòa tan trong đó sẽ được hấp thụ. Không giống như biểu bì, biểu bì không được bao phủ bởi lớp biểu bì và không có khí khổng. Dưới nó là vỏ não sơ cấp, bao gồm ngoại bì, trung bì và nội bì.
1 - các vùng đầu rễ; 2 - 6 - mặt cắt ngang ở các cấp khác nhau; Vks - xylem thứ cấp; Vf - phloem thứ cấp; K - cambium; Cô, metaxylem; Mf - ẩn dụ; Pd - biểu bì; Pks - protoxylem; Prd - quanh da; Pf - protofloem; PC - vòng tua máy; Рд - thân rễ; Ex - ngoại diên; En - nội bì; H - khởi động.
Các tế bào của ngoại bì gấp nếp dày đặc và có màng dày. Lớp vỏ ngoài đảm bảo dòng nước không đàn hồi vào các lớp sâu hơn của vỏ, đồng thời mang lại sức mạnh cho rễ từ bề mặt.
Lớp giữa của vỏ, trung bì, bao gồm các tế bào nhu mô thành mỏng và có cấu trúc lỏng lẻo do sự hiện diện của nhiều khoảng gian bào mà nước di chuyển đến ngoại bì.
Nội bì, hay lớp bên trong của vỏ não sơ cấp, bao gồm một hàng tế bào. Nó bao gồm các ô có vành đai Caspari và ô tiếp cận. Tế bào có đai Caspari có vỏ dày lên bên (xuyên tâm) và tiếp tuyến (cuối) đối diện với hình trụ trung tâm. Những hạt xenlulo này được ngâm tẩm với lignin và do đó không thể đi qua nước. Ngược lại, các tế bào dẫn truyền có thành mỏng và nằm đối diện với tia xylem. Nước đi vào bó dẫn hướng tâm của hình trụ trung tâm thông qua các ô dẫn.
Hình trụ trung tâm của rễ, hay tấm bia, bao gồm một số lớp tế bào. Một hoặc nhiều hàng tế bào chu kỳ nằm ngay dưới nội bì. Trong số này, trong quá trình chuyển sang cấu trúc thứ cấp của rễ ở cây hạt kín và cây hạt trần hai lá mầm, một cây giao thoa và phellogen (bần cambi) được hình thành. Ngoài ra, tế bào chu kỳ còn tham gia vào quá trình hình thành rễ bên. Các tế bào của chùm tia dẫn xuyên tâm nằm phía sau vòng tròn.
Bó dẫn được hình thành từ procambium. Đầu tiên, tế bào protofloem được hình thành, và sau đó là ở cấp độ của những sợi lông gốc đầu tiên - tế bào protoxylem. Tế bào của các mô dẫn phát sinh ngoại bào, tức là từ bề mặt của chùm tia, và tiếp tục phát triển theo hướng hướng tâm. Trong trường hợp này, các mạch đầu tiên, lớn hơn nằm ở trung tâm của tia xylem, và các mạch nhỏ hơn, có đường kính nhỏ hơn, nằm ở ngoại vi của tia xylem. Tế bào phloem nằm giữa các tia xylem.
Số lượng tia xylem phụ thuộc vào vị trí hệ thống của cây. Ví dụ, một số loài dương xỉ có thể chỉ có một tia xylem và một mảng phloem. Khi đó bó được gọi là quân vương. Đối với nhiều thực vật hạt kín hai lá mầm, các bó lưỡng bội có hai tia xylem là đặc trưng. Ngoài ra, họ còn có các cây có bó tri-, tứ và ngũ bội. Thực vật hạt kín một lá mầm được đặc trưng bởi các bó mạch đa phân với xylem nhiều tia.
Cấu trúc của chùm dẫn hướng tâm ảnh hưởng đến cách đặt các rễ bên. Trong rễ diarchic, chúng được hình thành giữa phloem và xylem, trong triarchic và tetrarchous, đối diện với xylem và trong polyarchic, đối diện với phloem.
Cấu trúc tế bào epiblema và vỏ não sơ cấp cung cấp áp suất nước cho rễ. Nước được hấp thụ bởi các lông rễ, từ đó nó đi vào màng tế bào của ngoại bì, sau đó đi vào các khoảng gian bào của trung bì, và từ chúng, qua các tế bào có vách mỏng, đi vào các mạch của chùm tia dẫn xuyên tâm. Vì các lông rễ có tổng bề mặt lớn hơn các tế bào thấm, nên tốc độ di chuyển của nước, và do đó áp lực của nó, tăng lên khi nó đến gần các mạch. Sự khác biệt trong các áp suất phát sinh là áp suất rễ, là một trong những cơ chế để nước chảy vào thân và các cơ quan khác của cây. Cơ chế quan trọng thứ hai cho sự chuyển động của nước là thoát hơi nước.
28.4.2. Cấu trúc giải phẫu thứ cấp của rễ
Ở thực vật hạt kín và thực vật hạt trần hai lá mầm, cấu trúc giải phẫu sơ cấp của rễ trong vùng dẫn truyền được bổ sung bởi các cấu trúc có nguồn gốc thứ cấp, được hình thành do sự xuất hiện và hoạt động mô phân sinh của các mô giáo dục bên thứ cấp - cambium và phellogen (bần cambium) (Hình . 3). Trong quá trình chuyển đổi sang cấu trúc thứ cấp, những thay đổi đáng kể sau đây xảy ra ở gốc.
A. Sự xuất hiện trong hình trụ trung tâm của rễ cambi và xylem thứ cấp và phloem thứ cấp do nó tạo ra, băng thông vượt xa các phần tử của chùm dẫn xuyên tâm ban đầu.
Các cơ quan khác nhau của thực vật bậc cao thực hiện các chức năng khác nhau. Vì vậy, rễ cây hấp thụ nước và khoáng chất, và quá trình quang hợp xảy ra ở lá, kết quả là chất hữu cơ được hình thành. Tuy nhiên, tất cả các tế bào thực vật đều cần cả nước và chất hữu cơ. Vì vậy, một hệ thống vận chuyển là cần thiết để đảm bảo cung cấp các chất cần thiết cho một số cơ quan từ những cơ quan khác. Ở thực vật (chủ yếu là thực vật hạt kín), chức năng này được thực hiện bởi mô dẫn điện.
Ở thực vật thân gỗ, các mô dẫn là một phần của gỗ và khốn nạn... Đối với gỗ được thực hiện hướng lên hiện tại: nước và chất khoáng bốc lên từ rễ. Bởi khốn thực hiện dòng điện đi xuống: có sự thoát ra của chất hữu cơ từ lá. Với tất cả những điều này, khái niệm "dòng điện đi lên" và "dòng điện đi xuống" không nên được hiểu theo nghĩa đen, như thể trong các mô dẫn nước luôn đi lên và các chất hữu cơ - đi xuống. Các chất có thể di chuyển theo chiều ngang và đôi khi theo hướng ngược lại. Ví dụ, chất hữu cơ đi đến chồi đang phát triển ở trên các mô dự trữ hoặc lá cây quang hợp.
Vì vậy, trong thực vật, sự chuyển động của dung dịch nước và chất hữu cơ được tách ra. Thành phần của gỗ bao gồm tàu thuyền, và trong tên khốn - ống sàng.
Mạch là một chuỗi các tế bào dài, chết. Một dung dịch nước di chuyển dọc theo chúng từ rễ. Nước dâng lên do áp suất của rễ và sự thoát hơi nước (bốc hơi nước từ lá). Thực vật hạt trần và dương xỉ thay vì mạch có khí quản, theo đó nước di chuyển chậm hơn. Do đó, các tàu có cấu trúc hoàn hảo hơn. Theo một cách khác, các tàu được gọi là khí quản.
Lý do tại sao nước trong các mạch chuyển động nhanh hơn trong khí quản nằm ở cấu trúc hơi khác nhau của chúng. Tế bào khí quản có nhiều lỗ xốp ở các điểm tiếp xúc với nhau (trên và dưới). Dung dịch nước được lọc qua các lỗ này. Mặt khác, mạch máu về bản chất là một ống rỗng, các tế bào của chúng có những lỗ lớn (lỗ thủng) ở phần tiếp giáp với nhau.
Các mạch này có nhiều độ dày khác nhau ở thành dọc của chúng. Điều này mang lại cho họ sức mạnh. Qua những nơi không có đặc, nước được vận chuyển theo phương ngang. Nó xâm nhập vào các tế bào của nhu mô và các mạch lân cận (các mạch thường được sắp xếp thành từng bó).
Các ống rây được hình thành bởi các tế bào sống kéo dài. Các chất hữu cơ di chuyển dọc theo chúng. Trên và dưới, các tế bào mạch máu được kết nối với nhau thông qua nhiều lỗ chân lông. Hợp chất này trông giống như một cái sàng, do đó có tên. Nó chỉ ra một chuỗi dài các tế bào. Mặc dù ống rây là tế bào sống nhưng chúng không có nhân và một số cấu trúc, bào quan khác cần thiết cho sự sống. Do đó, ống sàng có cái gọi là ô đồng hành hỗ trợ sinh kế của họ. Các ống đồng hành và ống được kết nối với nhau thông qua các lỗ rỗng đặc biệt.
Gỗ và libe không chỉ được làm bằng vải dẫn điện. Chúng cũng bao gồm nhu mô và các mô cơ học. Vải dẫn điện cùng với vải cơ học tạo thành bó sợi mạch máu... Nhu mô thường đóng vai trò mô dự trữ (nhất là ở gỗ).
Gỗ được gọi khác nhau xylem, và khốn - phloem.
Trong quá trình tiến hóa, đó là một trong những nguyên nhân giúp thực vật có thể sinh sống trên cạn. Trong bài viết của chúng tôi, chúng tôi sẽ xem xét các tính năng của cấu trúc và hoạt động của các phần tử của nó - ống sàng và bình.
Tính năng của vải dẫn điện
Khi những thay đổi lớn về điều kiện khí hậu xảy ra trên hành tinh, thực vật phải thích nghi với chúng. Trước đó, chúng đều sống hoàn toàn dưới nước. Trong môi trường không khí mặt đất, việc hút nước từ đất và vận chuyển đến tất cả các cơ quan của cây là cần thiết.
Có hai loại mô dẫn, các phần tử của chúng là mạch và ống rây:
- Bast, hoặc phloem, nằm gần bề mặt của thân cây hơn. Trên đó, các chất hữu cơ hình thành trong lá trong quá trình quang hợp sẽ di chuyển về phía rễ.
- Loại mô dẫn điện thứ hai được gọi là gỗ, hoặc xylem. Nó cung cấp một dòng điện tăng dần: từ gốc đến lá.
Ống rây của thực vật
Đây là những tế bào dẫn điện của con khốn. Chúng được ngăn cách với nhau bởi nhiều vách ngăn. Bề ngoài, cấu trúc của chúng giống như một cái sàng. Đây là nơi bắt nguồn của tên. Các ống rây của thực vật đang sống. Điều này là do áp suất dòng điện đi xuống yếu.
Các bức tường ngang của chúng bị xuyên thủng với một mạng lưới lỗ dày đặc. Và các tế bào chứa nhiều lỗ xuyên qua. Chúng đều là sinh vật nhân sơ. Điều này có nghĩa là chúng không có lõi chính thức hóa.
Các phần tử của tế bào chất của ống rây chỉ còn sống trên thời gian nhất định... Thời gian của giai đoạn này rất khác nhau - từ 2 đến 15 năm. Chỉ số này phụ thuộc vào loại cây và điều kiện sinh trưởng của nó. Các ống rây vận chuyển nước và chất hữu cơ được tổng hợp trong quá trình quang hợp từ lá xuống rễ.

Tàu
Không giống như các ống sàng, các phần tử này của mô dẫn điện là các tế bào chết. Nhìn bề ngoài, chúng giống hình ống. Tàu có màng dày đặc. Ở bên trong, chúng tạo thành những lớp dày trông giống như những chiếc vòng hoặc hình xoắn ốc.
Do cấu trúc này, các tàu có thể thực hiện chức năng của chúng. Nó bao gồm sự di chuyển của các dung dịch khoáng trong đất từ gốc đến lá.

Cơ chế dinh dưỡng của đất
Như vậy, trong cây đồng thời diễn ra sự chuyển động của các chất theo các hướng ngược nhau. Trong thực vật học, quá trình này được gọi là dòng điện tăng dần và giảm dần.
Nhưng lực nào làm cho nước từ đất chuyển động lên trên? Nó chỉ ra rằng điều này xảy ra dưới ảnh hưởng của áp suất rễ và sự thoát hơi nước - sự bốc hơi nước từ bề mặt của lá.
Đối với thực vật, quá trình này có ý nghĩa sống còn. Thực tế là chỉ trong đất mới có khoáng chất, nếu không có chất này thì sự phát triển của các mô và cơ quan sẽ không thể thực hiện được. Vì vậy, nitơ cần thiết cho sự phát triển của hệ thống rễ. Có rất nhiều nguyên tố này trong không khí - 75%. Nhưng thực vật không có khả năng cố định nitơ trong khí quyển, đó là lý do tại sao dinh dưỡng khoáng rất quan trọng đối với chúng.
Tăng lên, các phân tử nước bám chặt vào nhau và thành mạch máu. Trong trường hợp này, các lực phát sinh có thể nâng nước lên một độ cao khá - lên đến 140 m. Áp lực này buộc các dung dịch đất qua các lông rễ thấm vào vỏ cây và đến các mạch của xylem. Thông qua chúng, nước dâng lên thân cây. Hơn nữa, dưới ảnh hưởng của sự thoát hơi nước, nước sẽ đi vào lá.
Các ống rây cũng nằm trong các tĩnh mạch cạnh mạch. Các phần tử này mang dòng điện đi xuống. Dưới tầm ảnh hưởng ánh sáng mặt trời trong lục lạp của lá tổng hợp ra pôlipeptit. Thực vật dành chất hữu cơ này cho quá trình tăng trưởng và quan trọng.
Vì vậy, các mô dẫn điện của cây đảm bảo sự di chuyển của các dung dịch nước của các chất hữu cơ và khoáng chất trong cây. Các yếu tố cấu trúc của nó là các mạch và ống sàng.
Trong quá trình tiến hóa, với sự xuất hiện của các loài thực vật bậc cao trên cạn, chúng đã phát triển các mô đạt đến mức độ chuyên môn hóa cao nhất ở thực vật có hoa. Trong bài viết này, chúng ta sẽ xem xét chi tiết hơn mô thực vật là gì, chúng tồn tại ở những dạng nào, chức năng của chúng cũng như các đặc điểm cấu tạo của mô thực vật.
Vải gọi các nhóm ô có cấu trúc giống nhau và thực hiện các chức năng giống nhau.
Các mô thực vật chính được thể hiện trong hình bên dưới:
Các loại, chức năng và cấu trúc của các mô thực vật.


Các mô liên kết của thực vật.
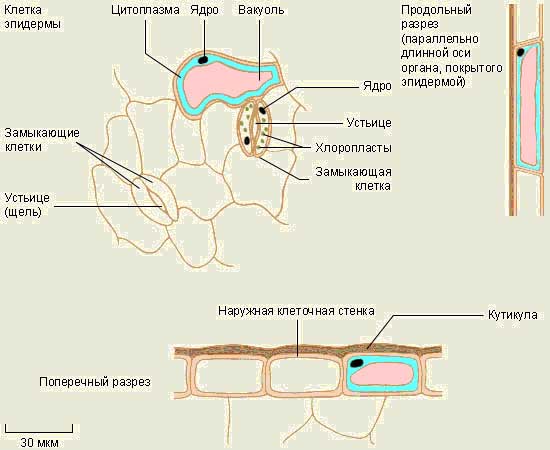

Bao bọc mô thực vật - lớp vỏ
Mô dẫn của thực vật.
| Tên vải | Kết cấu | Vị trí | Chức năng |
| 1. Tàu gỗ - xylem | Các ống rỗng với các bức tường nổi và các vật dụng đã chết | Gỗ (xylem) dọc theo gốc, thân, gân lá | Dẫn nước và khoáng từ đất lên rễ, thân, lá, hoa. |
| 2. ống sàng lớn - phloem Ô đồng hành hoặc ô đồng hành | Hàng dọc của tế bào sống có vách ngăn ngang sàng Tế bào chị em yếu tố sàngđã giữ lại cấu trúc của chúng | Bast (phloem), nằm dọc theo gốc, thân, gân lá Luôn nằm dọc theo các phần tử sàng (đi kèm với chúng) | Chuyển chất hữu cơ từ lá đến thân, rễ, hoa Tham gia tích cực đưa các chất hữu cơ qua ống rây của phloem. |
| 3. Tiến hành bó sợi mạch máu. | Một phức hợp của gỗ và gỗ tạp ở dạng các sợi cỏ riêng biệt và một khối cây vững chắc | Hình trụ trung tâm của rễ và thân; gân lá và hoa | Chở nước và khoáng trên gỗ; trên các chất hữu cơ; củng cố các cơ quan, kết nối của chúng thành một tổng thể duy nhất |



Mô cơ của thực vật.


Từ khóa » Bó Mạch Kín Gồm
-
TRƯỜng đẠi Học Nông Nghiệp Hà NỘi Bài Giảng Sinh HọC ĐẠi CưƠNG
-
BÀI GIẢNG THỰC VẬT HỌC - Tài Liệu Text - 123doc
-
Lớp Hành (Liliopsida)
-
Định Nghĩa Và Phân Loại Mô Dẫn - Dược Liệu
-
TV 2 Flashcards | Quizlet
-
Bó G Hình CH V, Bó M CH Kín | PDF - Scribd
-
Thực Vật Có Mạch – Wikipedia Tiếng Việt
-
Làm Thế Nào để Phân Biệt Cây Một Lá Mầm Và Cây Hai Lá Mầm?
-
Định Nghĩa Và Phân Loại Mô Dẫn | Thực Vật Dược Liệu
-
[PDF] MÔ THỰC VẬT - Trường Đại Học Nông Lâm TP.HCM
-
[PDF] Mô Phân Sinh Chu Cấp Ngành Hạt Trần Và Lớp Là Mạm Của Cay Hạt Tru
-
Mô Dẫn - VOER
-
[PDF] CƠ QUAN DINH DƯỠNG (Thân Cây)
-
Gãy Hai Xương Cẳng Chân - Health Việt Nam
-
[Tổng Hợp] Kiến Thức Y Khoa Về Giải Phẫu Dạ Dày Chi Tiết Nhất
-
Cơ Quan Sinh Dục Của Nữ Giới Gồm Những Gì? | TCI Hospital