하플로그룹 G-M201 - 요다위키
| 하플로그룹 G | |
|---|---|
 | |
| 발생가능시간 | 현재보다 4만8천5백 년 전에[1] |
| 가능한 원산지 | 서아시아[2] |
| 조상 | 하플로그룹 GHIJK |
| 후예 | 프라이머리: G1, G2 |
| 돌연변이의 정의 | M201, PF2957, L116, L154, L204, L240, L269, L402, L520, L521, L522, L523, L605, L769, L770, L836, L837, M201, P947/U6 페이지 |
하플로그룹 G(M201)는 인간 Y염색체 하플로그룹이다.HIZK는 부모 하플로그룹 GHIJK의 두 가지 분기 중 하나이며, 다른 하나는 HIZK입니다.
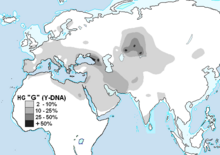
국가 인구 수준에서 G-M201은 조지아에서 가장 흔하게 발견되며, 코카서스의 오세티아인과 같은 많은 다른 지역 및 소수 집단에서 훨씬 더 높은 수준에서 발견된다.G-M201은 또한 유럽, 남아시아, 중앙아시아, 북아프리카의 소수 민족 집단에서 낮은 빈도로 널리 분포되어 있다.
가장 일반적으로 발생하는 서브레이드는 G1*(M285)와 G2(G-P287)의 많은 서브레이드이며, 특히 이전에는 G2a(P15), G2a1(G-FGC7535, 이전에는 G-L293, G2a2b2a(G-P303A)이다.5990 또는 G-Z1903)는 G2a3b1a3, G2b(G-M3115), G2b1(G-M377)은 G2b(구 G2b).
목차
- 1 오리진스
- 2 구조.
- 3 지리적 분포
- 3.1 선사 시대의 존재
- 3.2 모던한 존재감
- 4 G1(M285 또는 M342)
- 5 G2(P287)
- 5.1 G2a(P15)
- 5.2 G2a1(FGC7535)
- 5.3 G2a2a(PF3147)
- 5.4 G2a2b(L30, PF3267, S126, U8)
- 5.5 G2a2b1(M406)
- 5.6 G2a2b2(CTS2488)
- 5.7 G2a2b2a(G-P303)
- 5.8 G2a2b2b(PF3359)
- 5.9 G2b1(M377)
- 6 SNP의 정의
- 7 G-M201의 쟁쟁한 멤버
- 8 「 」를 참조해 주세요.
- 9 레퍼런스
- 10 외부 링크
오리진스
G-M201의 기원에 대해서는 다양한[2] 추정 날짜와 장소가 제시되고 있으며, 대부분은 서아시아에서 이루어지고 있다.
2012년, Siiri Rootsi 등의 논문은 "우리는 하플로그룹 G의 지리적 기원이 아나톨리아 동부, 아르메니아 또는 [3]이란 서부 근처에 있을 것으로 추정한다"고 제안했다.
이전에 내셔널 지오그래픽 소사이어티는 그 기원을 3만년 전 중동에 두고 하플로그룹을 들고 다녔던 사람들이 신석기 [4]시대의 확산에 참여했을 것으로 추정했다.
두 학술논문도 중동에서 유래를 제시하고 있지만 날짜는 다르다.세미노 등(2000년)은 17,000년 [5]전에 제안했습니다.Cinnioglu 등(2004)는 돌연변이가 불과 9,500년 [6]전에 일어났다고 주장했다.
히말라야 [7]산기슭에 가까운 지역에서 기원한 것으로 보이는 더 동쪽의 기원도 언급되고 있다.
구조.
G*(M201)
- G1(M285, M342)
- G1a(P20.1, P20.2, P20.3)
- G1a1(L201, L202, L203)
- G1b(L830, L831, L832, L834, L835)
- G1a(P20.1, P20.2, P20.3)
- G2(P287)
- G2a(P15, U5, L31/S149, L149)
- G2a1(L293^)
- G2a1a (P16.1, P16.2)
- G2a1a1(P18.1, P18.2, P18.3)
- G2a1a (P16.1, P16.2)
- G2a2(L223^)
- G2a2a(M286)
- G2a2b(L91)
- G2a2b1(L166, L167)
- G2a3(L30/S126, L32/S148, L190/M485)
- G2a3a(M406)
- G2a3a1(L14/Page57/S130/U16, L90/S133)
- G2a3a2(L645)
- G2a3b(L141.1)
- G2a3b1(P303/S135)
- G2a3b1a (L140)
- G2a3b1a1(U)
- G2a3b1a1a(L13/S131/U13, L78/M527)
- G2a3b1a1a1(L1263^)
- G2a3b1a1b(L1266^)
- G2a3b1a1b1(L1264^, L1265^, L1268^)
- G2a3b1a1a(L13/S131/U13, L78/M527)
- G2a3b1a2(L497, L353.1^, L353.2^)
- G2a3b1a2a(Z725^)
- G2a3b1a2a1(L43/S147)
- G2a3b1a2a1a(L42/S146)
- G2a3b1a2a1(L43/S147)
- G2a3b1a2a(Z725^)
- G2a3b1a3(Z1903^)
- G2a3b1a3a(Z724^)
- G2a3b1a3a1(L640)
- G2a3b1a3a(Z724^)
- G2a3b1a4(L660, L662)
- G2a3b1a1(U)
- G2a3b1b(L694)
- G2a3b1a (L140)
- G2a3b2(L177.1, L177.2, L177.3)
- G2a3b1(P303/S135)
- G2a3a(M406)
- G2a1(L293^)
- G2b(M377, L72, L183)
- G2b1(M283)
- G2a(P15, U5, L31/S149, L149)
(여기에 있는 하위 분류는 ISOGG가 사용하는 Y-DNA SNP 정의에 부합한다. 2012년에 연구 연구에서 한 남성에게만 발견된 여러 범주가 ISOG 트리에서 제거되어 일부 이름이 변경되었다.)
지리적 분포
선사 시대의 존재
하플로그룹 G2a(G-P15)의 염기서열을[self-published source?] 가진 고대 G-M201은 기원전 5000~3000년 사이의 신석기 시대 인류 유적에서 확인되었다.이 신석기 유럽인들은 아나톨리아에서 온 신석기 농부들의 후손으로, 세계에서 가장 먼저 농업을 시작한 사람들 중 일부입니다.G-M201은 기원전 8300-7600년의 본쿠클루와 기원전 [8][9]6419-6238년의 바르신 같은 신석기 아나톨리아 유적지에서도 발견되었다.
게다가 유럽 신석기 시대의 남성 유골의 대부분은 지금까지 이 하플로그룹에 속하는 Y-DNA를 생산해 왔다.고대 DNA 테스트에 의해 하플로그룹 G2a를 가지고 있는 것으로 확인된 가장 오래된 유골은 스페인 카탈로니아의 레스 플레인 도호스톨 근처의 아벨라네르 동굴 매장지에서 발견된 5개이며 기원전 5000년 [10]경의 방사성 탄소에 의해 확인되었다.
독일 작센안할트주 데렌부르크 메렌스티에그 2세로 알려진 신석기시대 공동묘지에서 발견된 유골은 G2a3(G-S126) 또는 하위 계단의 것으로 보인다.그것은 선형 밴드 세라믹 문화(LBK)의 매장 유물과 함께 발견되었다.이 유골은 방사성 탄소 연대 측정으로 연대를 알 수 없지만, 다른 유골들은 5,100년에서 6,100년 전으로 거슬러 올라간다.식별된 가장 상세한 SNP 변환은 G2a3를 [11]정의하는 S126(L30)입니다.
G2a는 또한 프랑스 남부 신석기 후기 농부들의 서식지인 트레유에서 약 5000년 [12]전의 고대 Y-DNA 샘플 22개 중 20개에서 발견되었다.같은 시기의 네 번째 유적지는 이탈리아 알프스의 외츠탈에서 얼음 사람 외치의 미라 유적이 발견되었다.아이스맨은 하플로그룹 G2a2b(이전의 G2a4)에 속합니다.
하플로그룹 G2a2b는 오늘날 유럽에서 보기 드문 그룹이다.스페인 연구의 저자들은 Avellaner 남성이 짧은 탠덤 반복(STR) 마커 테스트에서 희귀한 마커 값을 가지고 있음을 나타냈다.
모던한 존재감
주요 기사:국가별 하플로그룹 G(Y-DNA)러시아, 우크라이나 및 중앙아시아에서는 국가 수준의 평균 비율이 약 1% 이하임에도 불구하고 다양한 소수 민족 및/또는 특정 지역의 거주자가 세계에서 가장 높은 수준으로 G-M201을 보유하고 있다.카자흐스탄의 마자르족과 아르긴족(또는 씨족)은 현대 민족 중 가장 높은 수준의 G-M201을 가지고 있는 것으로 밝혀졌다.마자르족 중에서 G1은 87%의 비율로 발견되었다.아르긴족에 대한 별도의 연구는 남성의 71%가 G1에 속한다는 것을 알아냈다.러시아 북카프카스에서도 카바르디니아와 오세티안 인구는 G-M201의 높은 비율로 유명하다.북오세티야 디고라는 74%가 [14]G로 단일 도시에서 가장 높은 G 농도를 보였다.하플로그룹 G는 중국 북부와 같은 먼 동쪽 지역에서 발견되는데, G는 위구르족과 같은 소수 집단에서 더 실질적인 비율에 도달할 수 있다.
터키, 남카프카스, 이란에서는 하플로그룹 G가 전국 인구 중 가장 높은 비율을 차지하고 있다.터키 남성들 중 11%가 [6]G이다.이란에서는 하플로그룹 G가 전국 각지에서 인구의 13~15%에 이른다.바흐티아리족, 탈리시족, 길라키족, 마잔다라니족, 이란 아제리족에서 10% 이상 발견되는 반면, 이란 아랍인과 일부 [15]대도시에서는 5%에 가깝다.남부 코카서스 국가의 YHRD 데이터베이스 샘플 중 아바지니아에서 29%, 그루지야에서 31%, 아제르바이잔에서 2%, 아르메니아에서 18%가 G 샘플로 보인다.
흑해 서부의 유럽에서는, 하플로그룹 G는 대륙의 [16]대부분에서 평균 인구의 약 5%에서 발견된다.스칸디나비아, 구소련 최서단 공화국, 폴란드, 아이슬란드와 영국 제도에서 G 농도는 이 평균 이하로 떨어집니다.유럽 내에는 특이한 집중현상이 있는 것 같다.웨일즈에서는, 독특한 G2a3b1 타입(DYS388=13 및 DYS594=11)이 그곳을 지배하고 인구의 G 퍼센티지를 영국보다 높게 밀어낸다.
오스트리아 서부의 티롤(티롤)에서는 G-M201의 비율이 40% 이상에 달할 수 있습니다.아마 가장 유명한 예는 소위 "아이스만"인 외치의 고대 유적일 것입니다.이탈리아 서부의 사르디니아 섬의 북부 및 고지대에서는 G 퍼센티지가 인구의[17] 11%에 달하고, 또 다른 연구에서는 템피오 마을의 21%에 달했다.그리스 크레타 섬에서는 남성의 약 7%[18]에서 [19]11%가 하플로그룹 G에 속합니다.크로아티아 북동부의 오시예크 마을에서 G는 [20]남성의 14%에서 발견되었다.이 도시는 유럽의 또 다른 하플로그룹 G 초점 지역인 알프스 티롤/티롤 지역에서 시작되는 드라바 강둑에 있습니다.더 북쪽에서는 헝가리계 남성의 8%와 보헤미안(체코)계 남성의 5.1%가 하플로그룹 G에 속해 있는 것으로 밝혀졌다.
남아시아에서는 일부 소수 민족이 칼라쉬의 약 [21]18~20%,[22] 브라후이의 [22]약 16%, 파슈툰의 [21]약 11.5%의 농도로 하플로그룹 G를 보유하고 있지만 파키스탄 인구의 [21]약 3%만이 가지고 있다.936명의 인도인을 대상으로 한 연구에서, 하플로그룹 G는 표본의 1% 미만을 차지했고, 테스트된 북서부 인도 [23]인구에는 전혀 없었다.스리랑카 및 말레이시아에서 채취한 샘플의 약 6%가 하플로그룹 G로 보고되었지만,[24] 아시아에 있는 인도양 또는 태평양의 다른 연안 육지에서는 발견되지 않았다.하플로그룹 G-M201은 신석기 시대에는 상대적으로 존재하지 않았던 것으로 추정되며, 예를 들어 G2a-P15 서브레이드의 빈도는 인도 원주민 [25]개체군에서는 무시해도 될 정도였다.
중동에서는 거의 모든 지역에서 [26][27]하플로그룹 G가 인구의 약 3%를 차지한다.드루즈족 중 10%가 하플로그룹 [28]G인 것으로 밝혀졌다.
유대인 남성의 약 10%가 하플로그룹 [citation needed]G이다.
하플로그룹 G는 사하라 이남 아프리카나 아프리카 뿔 남쪽에서 원주민 개체군 중 드물게 발견된다.이집트에서는 G 퍼센티지가 2%에서 9%[29][30][31] 사이라는 연구 결과가 나왔다.북아프리카 베르베르인의 3%가 하플로그룹 [32]G이며, 아랍계 모로코인의 2%, 베르베르계 모로코인의 0.8%도 [33]G로 나타났다.
아메리카에서 하플로그룹 G의 비율은 구세계 국가에서 이민 온 사람들의 수에 해당한다.그것은 비 원주민과의 혼인이 발생한 경우를 제외하고는 아메리카 원주민들 사이에서 발견되지 않는다.그것은 멕시코 메스티소에서 발견되었다.
G1(M285 또는 M342)
주요 기사:하플로그룹 G1 (Y-DNA)거의 모든 하플로그룹 G1 사람은 짧은 탠덤 반복(STR) 마커 DYS392에서 12의 값을 가지며, 모두 이 그룹을 특징짓는 M285 또는 M342 SNP 돌연변이를 가진다.이 값 12는 G1 이외의 다른 G 범주에서 흔치 않다.
G1a, G1a1, G1b의 서브레이드가 존재한다.
단일 국가에서 G1과 그 하위 계층의 농도가 가장 높게 보고된 것은 이란이며, 다음으로 서쪽의 인접 국가에서 가장 많이 나타난다.
에는 독특한 아슈케나지 유대인과 카자흐 subclades STR 마커 값 조합에 기초한다.
G2(P287)
이 그룹지만 모두 G2subclades에 부정적인 속하남자들은haplogroup G명으로 이루어진 작은 수를 뜻한다.P287 애리조나 대학교의 창법으로 널리 2007년 말로 잘 알려 져 확인되었다.그것의 식별 G범주의 상당한 개명을 일으켰다.
G2a(P15)
이 그룹도 G2a subclades에 부정적인 속하하플로 그룹 G남자들, 유럽에서지만 터키의 지금까지 형편없이 시험 지역 동쪽으로 상당한 그룹을 대표할 것 흔하지 않다.P15 애리조나 대학교의 창법으로 널리 2002년에 의해 알려져 확인되었다.그것의 염색체 위치 21653414으로 등록되어.G2a 중세에서 발견되었다 Ergolding, 바이에른, 독일의 한7th-세기 CE에는 상류층 무덤에 있지만, G2a subclades 시험 받은 것이 아니라 남아 있다.[34]
지금까지 P15와 같은 커버리지를 가지는 복수의 SNP가 있습니다.Y염색소 로케이션을 수반하는 것은, U5(rs2178500), L149(8486380) 및 L31(일명 S149)입니다(r3561775).12538148).P15 돌연변이를 가진 남성이 이들 중 하나 또는 그 반대인 경우, 그 결과는 새로운 G2a 범주의 기초가 될 것이다.
G2a1(FGC7535)
주요 기사:하플로그룹 G-FGC7535하플로그룹 G2a1(G-FGC753 및 이전에 G-L293)과 그 하위 계층은 코카서스 산맥 일부 지역에서 하플로그룹 G 샘플의 대부분을 나타낸다.그들은 다른 곳에서는 아주 적은 수의 사람들만 볼 수 있다.지금까지의 모든 G2a1 사람은 STR 마커 DYS392에서 10의 값을 가지고 있다. G2a1a 사람은 일반적으로 대부분의 G 사람에게서 볼 수 있는 것보다 16, 17 또는 18과 같은 DYS385b에 대해 더 높은 값을 가지고 있다.
러시아 중북부 코카서스 지역의 북오세티안은 이용 가능한 샘플에 근거해 압도적으로 G2a1 하위군에 속한다.북오세티야 남쪽의 남오세티야와 스반에는 G2a1 인구가 상당히 많지만 인구 비율은 아직 정해지지 않았다.
SNP P18 돌연변이의 존재는 G2a1a의 유일한 서브레이드인 G2a1a를 특징짓는다.P16과 P18의 신뢰성에 의문이 제기되어 SNP의 개별 컴포넌트를 조사해야 합니다.
북동유럽 혈통을 가진 Ashkenazi 유대인 G2a1a 남성은 STR 마커 값을 바탕으로 뚜렷한 집단을 형성한다.코카서스에서 온 남자들과 동유럽에서 온 남자들도 독특한 STR 클러스터를 형성한다.
G2a2a(PF3147)
G-PF3147(이전의 G-L223 및 G-PF3146)은, L223 변이를 가지는 것이 특징입니다.L223은 Y 염색체 rs810801과 6405148에서 C에서 G로의 돌연변이와 함께 발견된다. L223은 2009년 23andMe의 표본에서 처음 확인되었지만 개별 테스트로서 문제가 있음이 입증되었으며, 첫 성공적인 결과는 2011년 말 할당된 L223 라벨로 패밀리 트리 DNA에서 보고되었다.그 후 다음과 같은 여러 하위 분류군이 L223에 속한다는 것을 알게 되었다.
G-L91은 2009년에 식별되었다.유럽 알프스 산맥에서 최소 5,000년 전에 사망한 아이스맨 "외치"가 그 구성원이다.G-L91은 G-L91에 속하는 남성의 상당 부분을 포함하는 것으로 보인다. L91은 지금까지 유럽과 북아프리카의 산재된 지역과 아르메니아에서 발견된다.G-L91에는 STR 마커 DYS19에 대한 이중 값을 가진 남성들이 포함되지만, L91+가 아닌 G2a2 남성들도 있다.G2a1과 G2a3 서브레이드에서는 이중 19 값 상황은 보이지 않는다.L91 돌연변이는 Y염색체의 21327383과 rs35474563에서 발견된다.정방향 프라이머는 입니다.정방향 프라이머는 입니다.돌연변이는 C에서 [citation needed]T로의 변화를 수반한다. L223은 rs13304806의 Y 염색체에서 발견된다.
G-M286 서브레이드(M286+)는 G-L91에 비해 작다. 표본은 영국, 독일, 보스니아, 스페인, 키프로스, 터키, 아르메니아, 조지아, 레바논, 시리아 및 오만에서 확인되었다.영국 검체는 대부분의 경우 STR 마커 DYS19에 대해 일관되지 않은 이중 값을 가집니다.M286은 Stanford University에서 염색체 위치 21151187에서 처음 확인되었으며 G에서 A로의 돌연변이입니다.
세 번째 서브레이드를 특징짓는 L293 SNP는 2010년 6월 패밀리 트리 DNA에서 확인되었다.그것은 소수 히스패닉 남성들로 구성되어 있으며, 그들은 모두 YCA 마커의 13,21이라는 이상한 가치를 가지고 있다.돌연변이는 Y염색체 10595022에서 발견되며 G에서 C로의 변화이다.
G2a2b(L30, PF3267, S126, U8)
G-L30(또한 G-PF3267, G-S126 또는 G-U8; G2a2b, 이전에는 G2a3) 이 그룹에 속하지만 모든 서브레이드에 음성인 남성은 오늘날 소수의 숫자를 나타낸다.이 하플로그룹은 독일 데렌부르크 미렌스티에그 2세 공동묘지에서 기원전 5000년경 신석기 시대의 골격에서 발견되었는데, 독일어로 선형 반데라믹(LBK)[11]으로 알려진 선형 도자기 문화의 일부를 형성하지만 G2a3 하위 계층에 대해서는 테스트되지 않았습니다.
G2a2b1(M406)
주요 기사:하플로그룹 G-M406G-M406*(G2a2b1*; 이전 G2a3a*)와 그 서브레이드는 터키와 지중해 동부 해안 지역에서 가장 흔하게 볼 수 있으며, 모든 제조사의 최대 5%, 하플로그룹 G 샘플의 50%를 구성할 수 있다.G2a2b1은 북유럽보다 남유럽에서 더 흔하다.이탈리아를 제외한 유럽에서는 G2a2b1이 G 샘플의 20% 미만을 구성한다.G2a2b1은 지금까지 북아프리카나 남아시아에서는 거의 표면화되지 않았지만 코카서스 산맥 지역과 이란에서는 G 인구의 작은 비율을 차지한다.
STR 마커 DYS390에서 G2a2b1의 비교적 높은 비율은 21이다.DYS391 마커는 G2a2b1 사람에서 대부분 10의 값을 가지지만 때로는 11의 값을 가지며, DYS392는 거의 항상 11입니다.샘플이 이 세 마커에 대해 표시된 기준을 충족하면 샘플은 G2a2b1일 가능성이 높습니다.
G2a2b2(CTS2488)
G-CTS2488 또는 G2a2b2(일명 G-L141.1, 이전에는 G-141 및 G2a3b)는 2009년 중반에만 패밀리 트리 DNA에서 확인되었다.거의 모든 L141 남자들은 L141 하위 계단에 속해 있다.영국 제도, 시칠리아, 터키 혈통을 가진 사람들의 샘플이 확인되었다.지금까지 L141 서브레이드에 속하지 않은 L141 사람은 STR 마커 DYS490에서 11의 값을 가진다. 이는 다른 G 범주에서 드문 발견이다.L141 돌연변이는 Y 염색체 2948607에서 발견된다.L141 돌연변이는 [35]삽입을 수반합니다.
G2a2b2a(G-P303)
주요 기사:하플로그룹 G-P303G2a2b2a*(이전의 G2a3b1*)로도 알려진 G-P303*와 그 서브레이드는 현재 러시아 남부와 코카서스 산맥에 집중되어 있으며, 하층에는 유럽과 서남아시아의 다른 지역, 특히 터키, 이란, 중동에서 G2A2가 발생할 가능성이 있는 지역에 집중되어 있다.G2a2b2a는 인도에서도 발견된다.
G-P303 멤버의 대부분은 G-P303이 아닌 하위 분류군 중 하나에 속합니다.*
사용 가능한 샘플에 기초한 가장 큰 G-P303* 하위 계통은 STR 마커 DYS388에서 거의 모든 사람이 13의 값을 갖는 것이다.SNP L497은 이러한 남성들을 포함하지만, 대부분의 G-L497 남성들은 G-DYS388=13으로 알려진 하위 계층 G-Z725에 속해 있다.특정 SNP 또는 흔치 않은 STR 마커 이상 유무를 특징으로 하는 DYS388=13 남성의 추가 서브레이드가 있다.이 그룹의 구성원들은 유럽과 [3]중동에서 발견되었다.
다음으로 큰 G-P303 서브레이드는 U1 돌연변이의 존재로 특징지어진다.그러나 U1 남성의 높은 비율은 G-L13/S13과 Z1266(G2a3b1a1b) 두 개의 하위 계단에 속합니다.G-L13 서브레이드는 중앙 북유럽에서 가장 흔하며, G-Z1266은 서부 코카서스 산맥에서 가장 흔하다.
최종 주요 서브레이드는 SNP Z1903의 존재와 마커 DYS568에서 9의 값을 갖는 것이 특징이다.G-Z1903 남성의 높은 비율이 하위 계층인 G-Z724에 속합니다.
지금까지 기술된 개별 인구 중 G-P303의 가장 높은 비율은 스페인 동부 해안의 이비자 섬에 있다.이 집단은 스페인 종교재판 기간 동안 섬으로 도망쳤던 암호 유대인들과 관련이 있으며, 이 중 상당 부분은 G-Z725(DYS388=13)[36]로 확인된다.
G2a2b2b(PF3359)
G-PF3359(또는 G2a2b2b, 이전 G2a3b2)는 2013년 이전에는 G-L177로 알려져 있었다.SNP L177(일명.k.a. L1771.1/ L177_1, L1771.2/ L177_2, L177.3/ L177_3)은 "신뢰할 수 없는 회문 snp"[2][37]로 판명된 후 2013년에 ISOG에 의해 식별자로 철회되었다.
G-PF3359로 확인된 고대 DNA는 현재(BP), 헝가리(G-F1193*), 스페인([2]G-PF3359*) 4700의 고고학 유적지에서 발견되었다.
G-PF3359의 구성원은 G-P303에 포함된 남성보다 수가 적을 수 있지만, 관련 돌연변이에 대한 테스트는 소량만 수행되었다.지금까지 이에 긍정적인 남자들은 아일랜드, 영국, 네덜란드, 레바논, 터키(아르메니아 성) 혈통을 가지고 있었다.공유 STR 마커를 기반으로 한 여러 G-PF3359 서브레이드가 존재할 수 있습니다.
이 그룹에서 남성을 구분하는 STR 마커 값의 수는 [38][self-published source?]관련 남성의 수가 적음에도 불구하고 G-PF3359가 비교적 오래된 그룹임을 시사한다.관련된 돌연변이는 복잡하고 해석하기 어려울 수 있다.
G2b1(M377)
주요 기사:하플로 그룹 G-M377(Y-DNA)G-M377는, 현재는 G2b1로 알려진, 이전에 지정되었다 G2b과 G2c.3 다른 G2b haplotypes 현재까지 확인되는 등 밀접하게 관련된 아슈케나지 유대인들의 clade을 대표하는 사실상 모든 G2b명:한 터크 카르스에서 북동쪽 터키에서 아르메니아 근처에, 한 파슈툰, 그리고 하나의 Burusho 파키스탄에서.
G-M377의 파키스탄 북부에 위치한 극단적인 보기 드문 존재 이 곳에 산다고 G2b의 원본 지구 밖에 있고 그곳에서 역사적 기간에서, 더 서쪽(둘 다 아케메 네스 페르시아 제국, 알렉산더 대왕에 의해 정복되며 파키스탄 의 부분입니다, 그리고 그 후 그리스-박트리아 왕국의 부분을 차지하고)아마도에서 데려왔다 나타날 수도 있다.이 두 파키스탄 G-M377 haplotypes는 아슈케나지 유대인 clade 오며, 그래서 전혀의 공통된 기원을 표시하지 않는다 다른 있다고 보도했다.터키인 G-M377지만 동일하지 가깝다.만약 시험 다른 인구 집단에서 — 이 있다는 G-M377 근동 지역의 낮은 수준에서 발생한다 G-M377 haplotypes을 드러낼 지 지켜볼 일이다.
지금까지 테스트된 모든 G-M377 남성은 다른 대부분의 G 하플로타입에서 아직 발견되지 않은 RecLOH 이벤트의 결과인 DYS425 마커(DYS371 회문형 STR의 누락된 "T" 대립 유전자)에 대한 희귀한 null 값을 가지고 있다.세계 여러 지역에서 온 이스라엘 내 유대인들 중 G-M377은 [39]한 연구에서 3.7%를 차지했다.
하플로그룹 G-M377은 아프가니스탄 와르다크 지역에서 파슈툰족 5명의 표본 중 60%의 빈도로 발견되었다.이는 로컬 설립자 [40]효과 때문일 수 있습니다.
SNP의 정의
국제유전자계보학회(ISOGG)는 [41]하플로그룹 카테고리의 최신 컨센서스 버전을 유지하고 있다.이러한 분류는 공유 SNP 돌연변이를 기반으로 합니다.새로운 SNP가 검출되면 새로운 이름이 하플로그룹 카테고리에 할당될 수 있습니다.카테고리에 대한 대대적인 수정이 이루어진 2008년까지 G 카테고리는 거의 없었다.2009년부터 2012년까지 더 많은 G SNP가 식별되어 더 많은 변경이 이루어졌습니다.2008년까지 애리조나 대학(P 명칭), 스탠포드 대학(M 명칭) 또는 센트럴 플로리다 대학(U 명칭)의 연구소에서 새로운 G SNP가 보고되었습니다.2008년부터 패밀리 트리 DNA(L 명칭) 및 Ethnoancestry(S 명칭)에서 추가 G SNP가 식별되었다.후자의 연구실에서는 23andMe에서 약 2,000개의 SNP에 대해 테스트한 개인이 보고한 원시 데이터 결과를 사용하여 새로운 L 또는 S 지정 SNP 테스트를 실시했습니다.2009-10년에 특정 Y염색체 세그먼트의 염기서열을 지정하는 패밀리 트리 DNA의 Y프로젝트 워크 스루(Walk through the Y Project)는 L 명칭의 새로운 G SNP를 다수 제공했다.2012년부터 1000개의 게놈 프로젝트 데이터에서 시민 연구자가 최초로 확인한 Z 명칭의 SNP가 등장하기 시작했다.
SNP는 가장 신뢰할 수 있는 분류 방법을 제공하므로 각각은 공식 G 카테고리를 나타낼 수 있습니다.카테고리에는 문자와 숫자가 번갈아 표시됩니다.그러나 짧은 탠덤 반복 마커(STR)에서 발견되는 비정상적인 값 또는 비정상적인 값 조합도 추가 분류의 기초를 제공할 수 있다.새로운 SNP를 식별하려면 하나 이상의 카테고리를 변경해야 합니다.
하플로그룹 G를 특징짓는 M201 SNP 돌연변이는 스탠포드 대학에서 확인되었으며 [42]2001년에 처음 보고되었습니다.M201의 기술사양은 다음과 같습니다.refSNPid is rs2032636.....13536923의 Y염색체 위치는 ....앞으로 프라이머는 ....역 프라이머는 ....G에서 T로의 변화를 수반한다.
많은 SNP가 M201과 동일한 커버리지를 가진 것으로 확인되었습니다.M201이 먼저 확인되었기 때문에 G 사람에 대한 테스트 시 사용되는 표준 SNP 테스트입니다.이러한 대체 SNP 중 하나가 M201의 하위 계열을 나타내는지 여부를 판단하기 위해, 대체 SNP는 알려진 G의 하위 계단에 대해 음성인 G 사람을 대상으로 테스트해야 한다.그러한 범주에 속하는 사람은 극히 소수이며, M201을 제외한 소수의 사람들만이 G 등가 SNP 검사를 받았다.
다음 SNP는 지금까지 M201 등가물로 식별되었습니다.L116, L154, L269, L294, L240, P257, L402, L520, L521, L522, L523, L605, 페이지94, U2, U7, U6L240은 [44]2009년에 식별되었습니다."U" SNP는 2006년에 확인되었지만 [45]2009년에야 발표되었습니다.
또한 M201과 같은 커버리지를 가진 것으로 생각되는 다른 여러 SNP가 있습니다.Y 염색체의 rs9786910, rs9786537, rs2713254, rs35567891 및 rs34621155에서 찾을 수 있습니다.아직 어떤 연구실도 속기명을 부여하지 않았어요
G-M201의 쟁쟁한 멤버
다음 항목도 참조하십시오.역사적 인물로부터 도출된 유전자 결과 목록"외치"로 알려진 그 남자는 오스트리아와 이탈리아 사이의 현대 국경에서 발견된 기원전 4천 년기의 유골은 G-L91 (G2a2b)의 일원이다.
14세기 후반이나 15세기 초에 스웨덴에서 태어난 개인의 혈통을 추적하는 일족인 이른바 부레족(Bure kinship)의 많은 구성원들은 G2a2b2a1a1a1a로 알려진 G-Y12970(G-Y12971과 G-Z30735)에 속해 있는 것으로 알려졌다.
2012년 발굴된 영국의 리처드 3세는 G2(G-P287)[46] 멤버였다.
조셉 스탈린(본명 Ioseb Besarionis dz jug Jugashvili)은 그의 손자 중 한 명(Allexander Burdonsky)의 유전자 검사 결과, Y-DNA 하플로그룹 G2a1a(FGC595/Z653)의 [2][47]일원이었다.하플로그룹 G에 속하는 다른 수컷은 다음과 같다.독일계 미국인 개척과 병사 필립 Hamman, 필립의 5일 위대한 손자 셰인 Hamman 미국 올림픽 weightlifter,[48]미국 정치인 Linn은 은행들은 물리학자 존 G크레이머와 미국인 배우 제임스 프랜시스 커스, 제이크 질 랜홀, 미국 전 프로 농구 선수 래리 버드, 미국인 사업가 Najeeb Halaby, 미국 변호사와 administ.쥐.또는 뉴턴 미노, 미국 변호사이자 독립선언서 서명자인 리처드 [49][self-published source?]스톡턴입니다.
알 카포네는 미국의 갱스터이자 사업가였으며, Geni.com에 따르면 Y-DNA 하플로그룹 [50][self-published source?]G-P303의 멤버였다.
유야는 투탕카멘의 증조부였다.고대 Y-STR 프로파일에 기초한 하플로타입 G2a 예측.유야는 아멘호테프 3세의 주요 고문으로 '왕의 부관', '말의 주인' 등의 직책을 맡았으며, 그의 직함인 '신의 아버지'는 그가 아멘호테프의 장인이라는 것을 구체적으로 언급했을 것이다.그의 고향인 아크민에서, 유야는 이 지역의 "신"인 민의 예언자였고, 이 신의 "소 [51]관리"를 지냈다.
「 」를 참조해 주세요.
- 하플로그룹 G를 가진 유대인
- 유럽의 유전사
- 유전계보
- 세계 인구의 Y염색체 하플로그룹
- 코카서스인
- 막달레니아어
- 유럽의 Y-DNA 하플로그룹
- 코카서스 인구의 Y-DNA 하플로그룹
- 근동의 Y-DNA 하플로그룹
- 북아프리카의 Y-DNA 하플로그룹
- 민족별 Y-DNA 하플로그룹
레퍼런스
- ^ "G YTree".
- ^ a b c d e f ISOGG, 2017, Y-DNA 하플로그룹 G 및 하위 그룹 - 2017(2017년 5월 17일).
- ^ a b Rootsi, Siiri; Myres, Natalie M; Lin, Alice A; Järve, Mari; King, Roy J; Kutuev, Ildus; Cabrera, Vicente M; Khusnutdinova, Elza K; Varendi, Kärt; Sahakyan, Hovhannes; Behar, Doron M; Khusainova, Rita; Balanovsky, Oleg; Balanovska, Elena; Rudan, Pavao; Yepiskoposyan, Levon; Bahmanimehr, Ardeshir; Farjadian, Shirin; Kushniarevich, Alena; Herrera, Rene J; Grugni, Viola; Battaglia, Vincenza; Nici, Carmela; Crobu, Francesca; Karachanak, Sena; Kashani, Baharak Hooshiar; Houshmand, Massoud; Sanati, Mohammad H; Toncheva, Draga; Lisa, Antonella; Semino, Ornella; Chiaroni, Jacques; Cristofaro, Julie Di; Villems, Richard; Kivisild, Toomas; Underhill, Peter A (16 May 2012). "Distinguishing the co-ancestries of haplogroup G Y-chromosomes in the populations of Europe and the Caucasus". European Journal of Human Genetics. 20 (12): 1275–1282. doi:10.1038/ejhg.2012.86. PMC 3499744. PMID 22588667.
- ^ 인간 여정 지도: Happlogroup G(M201) 웨이백 머신에 2011-02-05 아카이브 완료
- ^ Semino O, Passarino G, Oefner PJ, Lin AA, Arbuzova S, Beckman LE, De Benedictis G, Francalacci P, Kouvatsi A, Limborska S, Marcikiae M, Mika A, Mika B, Primorac D, Santachiara-Benerecetti AS, Cavalli-Sforza LL, Underhill PA (November 2000). "The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective". Science. 290 (5494): 1155–9. Bibcode:2000Sci...290.1155S. doi:10.1126/science.290.5494.1155. PMID 11073453.
- ^ a b Cinnioğlu C, et al. (January 2004). "Excavating Y-chromosome haplotype strata in Anatolia". Human Genetics. 114 (2): 127–48. doi:10.1007/s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- ^ Mahal, David G.; Matsoukas, Ianis G. (23 January 2018). "The Geographic Origins of Ethnic Groups in the Indian Subcontinent: Exploring Ancient Footprints with Y-DNA Haplogroups". Frontiers in Genetics. 9: 4. doi:10.3389/fgene.2018.00004. PMC 5787057. PMID 29410676.
- ^ Feldman, Michal; Fernández-Domínguez, Eva; Reynolds, Luke; Baird, Douglas; Pearson, Jessica; Hershkovitz, Israel; May, Hila; Goring-Morris, Nigel; Benz, Marion; Gresky, Julia; Bianco, Raffaela A.; Fairbairn, Andrew; Mustafaoğlu, Gökhan; Stockhammer, Philipp W.; Posth, Cosimo; Haak, Wolfgang; Jeong, Choongwon; Krause, Johannes (19 March 2019). "Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia". Nature Communications. 10 (1): 1218. Bibcode:2019NatCo..10.1218F. doi:10.1038/s41467-019-09209-7. PMC 6425003. PMID 30890703. S2CID 83464300.
- ^ Hofmanová, Zuzana; Kreutzer, Susanne; Hellenthal, Garrett; Sell, Christian; Diekmann, Yoan; Díez-del-Molino, David; Dorp, Lucy van; López, Saioa; Kousathanas, Athanasios; Link, Vivian; Kirsanow, Karola; Cassidy, Lara M.; Martiniano, Rui; Strobel, Melanie; Scheu, Amelie; Kotsakis, Kostas; Halstead, Paul; Triantaphyllou, Sevi; Kyparissi-Apostolika, Nina; Urem-Kotsou, Dushanka-Christina; Ziota, Christina; Adaktylou, Fotini; Gopalan, Shyamalika; Bobo, Dean M.; Winkelbach, Laura; Blöcher, Jens; Unterländer, Martina; Leuenberger, Christoph; Çilingiroğlu, Çiler; Horejs, Barbara; Gerritsen, Fokke; Shennan, Stephen; Bradley, Daniel G.; Currat, Mathias; Veeramah, Krishna R.; Wegmann, Daniel; Thomas, Mark G.; Papageorgopoulou, Christina; Burger, Joachim (25 November 2015). "Early farmers from across Europe directly descended from Neolithic Aegeans" (PDF). bioRxiv: 032763. doi:10.1101/032763. S2CID 196682085.
- ^ Lacan, Marie; Keyser, Christine; Ricaut, François-Xavier; Brucato, Nicolas; Tarrús, Josep; Bosch, Angel; Guilaine, Jean; Crubézy, Eric; Ludes, Bertrand (8 November 2011). "Ancient DNA suggests the leading role played by men in the Neolithic dissemination". Proceedings of the National Academy of Sciences of the United States of America. 108 (45): 18255–18259. Bibcode:2011PNAS..10818255L. doi:10.1073/pnas.1113061108. PMC 3215063. PMID 22042855.
- ^ a b Haak W, Balanovsky O, Sanchez JJ, Koshel S, Zaporozhchenko V, Adler CJ, Der Sarkissian CS, Brandt G, Schwarz C, Nicklisch N, Dresely V, Fritsch B, Balanovska E, Villems R, Meller H, Alt KW, Cooper A (2010). Penny D (ed.). "Ancient DNA from European Early Neolithic Farmers Reveals Their Near Eastern Affinities". PLOS Biology. 8 (11): e1000536. doi:10.1371/journal.pbio.1000536. PMC 2976717. PMID 21085689.
- ^ Marie Lacan, Christine Keyser, Francis-Xavier Ricaut, Nicolas Brucato, Francis Duranthon, Jean Guilaine, Eric Crubézy 및 Bertrand Ludes, 고대 DNA는 신석기 지중해 루트를 통한 남성 확산, 2011년 5월 31일 전 미국 국립과학아카데미 온라인 인쇄되었습니다.
- ^ 보젠에 있는 고고학 박물관의 연구 프로젝트 책임자이자 보조인 에두아르트 에가터 비글 박사를 인터뷰합니다.Document-Movie: Christine Sprachmann의 "Hötzi, ein Archéologiekrimi" (예츠, 고고학 범죄)TV-Broadcast 2011년 8월 10일, br-alpha 2011년 9월 13일
- ^ Nasidze, Ivan; Sarkisian, Tamara; Kerimov, Azer; Stoneking, Mark (14 December 2002). "Testing hypotheses of language replacement in the Caucasus: evidence from the Y-chromosome". Human Genetics. 112 (3): 255–261. doi:10.1007/s00439-002-0874-4. PMID 12596050. S2CID 13232436.
- ^ Nasidze I, et al. (2008). "Close Genetic Relationship Between Semitic-speaking and Indo-European-speaking Groups in Iran". Annals of Human Genetics. 72 (Pt 2): 241–52. doi:10.1111/j.1469-1809.2007.00413.x. PMID 18205892. S2CID 5873833.
- ^ "G-Z7941 - Haplogroup G Project". google.com.
- ^ Zei G, et al. (2003). "From surnames to the history of Y chromosomes: the Sardinian population as a paradigm". European Journal of Human Genetics. 11 (10): 802–07. doi:10.1038/sj.ejhg.5201040. PMID 14512971.
- ^ Martinez L, et al. (April 2007). "Paleolithic Y-haplogroup heritage predominates in a Cretan highland plateau". European Journal of Human Genetics. 15 (4): 485–93. doi:10.1038/sj.ejhg.5201769. PMID 17264870.
- ^ King RJ, et al. (March 2008). "Differential Y-chromosome Anatolian influences on the Greek and Cretan Neolithic". Annals of Human Genetics. 72 (Pt 2): 205–14. doi:10.1111/j.1469-1809.2007.00414.x. PMID 18269686. S2CID 22406638.
- ^ Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri; Marjanovic, Damir; Primorac, Dragan; Hadziselimovic, Rifat; Vidovic, Stojko; Drobnic, Katia; Durmishi, Naser; Torroni, Antonio; Santachiara-Benerecetti, A Silvana; Underhill, Peter A; Semino, Ornella (24 December 2008). "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe". European Journal of Human Genetics. 17 (6): 820–830. doi:10.1038/ejhg.2008.249. PMC 2947100. PMID 19107149.
- ^ a b c Firasat S, Khaliq S, Mohyuddin A, Papaioannou M, Tyler-Smith C, Underhill PA, Ayub Q (2006). "Y Chromosomal Evidence for a Limited Greek Contribution to the Pathan Population of Pakistan". European Journal of Human Genetics. 15 (1): 121–26. doi:10.1038/sj.ejhg.5201726. PMC 2588664. PMID 17047675.
- ^ a b Sengupta S, et al. (February 2006). "Polarity and temporality of high-resolution y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists". American Journal of Human Genetics. 78 (2): 202–21. doi:10.1086/499411. PMC 1380230. PMID 16400607.
- ^ Sahoo, Sanghamitra; Singh, Anamika; Himabindu, G.; Banerjee, Jheelam; Sitalaximi, T.; Gaikwad, Sonali; Trivedi, R.; Endicott, Phillip; Kivisild, Toomas; Metspalu, Mait; Villems, Richard (2006-01-24). "A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios". Proceedings of the National Academy of Sciences. 103 (4): 843–848. Bibcode:2006PNAS..103..843S. doi:10.1073/pnas.0507714103. ISSN 0027-8424. PMC 1347984. PMID 16415161.
- ^ Hammer MF, et al. (2006). "Dual Origins of the Japanese: Common Ground for Hunter-Gatherer and Farmer Y-Chromosomes". Journal of Human Genetics. 51 (1): 47–58. doi:10.1007/s10038-005-0322-0. PMID 16328082.
- ^ Singh, Sakshi; Singh, Ashish; Rajkumar, Raja; Sampath Kumar, Katakam; Kadarkarai Samy, Subburaj; Nizamuddin, Sheikh; Singh, Amita; Ahmed Sheikh, Shahnawaz; Peddada, Vidya; Khanna, Vinee; Veeraiah, Pandichelvam; Pandit, Aridaman; Chaubey, Gyaneshwer; Singh, Lalji; Thangaraj, Kumarasamy (2016). "Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup". Scientific Reports. 6: 19157. Bibcode:2016NatSR...619157S. doi:10.1038/srep19157. PMC 4709632. PMID 26754573.
- ^ 그러나 레바논에서는 G가 인구의 6.5%, 이란에서는 약 10%를 차지한다.Flores C, et al. (2005). "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan". Journal of Human Genetics. 50 (9): 435–41. doi:10.1007/s10038-005-0274-4. PMID 16142507.
- ^ Cadenas AM, et al. (2008). "Chromosome Diversity Characterizes the Gulf of Oman". European Journal of Human Genetics. 16 (3): 374–86. doi:10.1038/sj.ejhg.5201934. PMID 17928816.
- ^ Shlush LI, et al. (2008). Gemmell NJ (ed.). "The Druze: A Population Genetic Refugium of the Near East". PLOS ONE. 3 (5): e2105. Bibcode:2008PLoSO...3.2105S. doi:10.1371/journal.pone.0002105. PMC 2324201. PMID 18461126.
- ^ European Journalof Human Genetics (2005) 13, 867–876 (cf) "아프리카의 Y 염색체와 mtDNA 변이의 대조적 패턴: 성편향 인구학적 과정의 증거"부록 A: Y 염색체 하플로타입 주파수)
- ^ Luis JR, et al. (2004). "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations". American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- ^ El-Sibai M, Platt DE, Haber M, Xue Y, Youhanna SC, Wells RS, Izaabel H, Sanyoura MF, Harmanani H, Bonab MA, Behbehani J, Hashwa F, Tyler-Smith C, Zalloua PA (2009). "Geographical Structure of the Y-Chromosomal Genetic Landscape of the Levant: A Coastal-Inland Contrast". Annals of Human Genetics. 73 (Pt6): 561–81. doi:10.1111/j.1469-1809.2009.00538.x. PMC 3312577. PMID 19686289.
- ^ Alonso S, Flores C, Cabrera V, Alonso A, Martín P, Albarrán C, Izagirre N, de la Rúa C, García O (2005). "The place of the Basques in the European Y-chromosome diversity landscape". European Journal of Human Genetics. 13 (12): 1293–1302. doi:10.1038/sj.ejhg.5201482. PMID 16094307.
- ^ Cruciani F, et al. (2002). "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". American Journal of Human Genetics. 70 (5): 1197–1214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- ^ Vanek D, Saskova L, Koch H (2009). "Kinship and Y-Chromosome Analysis of 7th Century Human Remains: Novel DNA Extraction and Typing Procedure for Ancient Material". Croatian Medical Journal. 50 (3): 286–95. doi:10.3325/cmj.2009.50.286. PMC 2702742. PMID 19480023.
- ^ ymap.ftdna.com[영구 데드링크]
- ^ Adams SM, Bosch E, Balaresque PL, Ballereau SJ, Lee AC, Arroyo E, López-Parra AM, Aler M, Grifo MS, Brion M, Carracedo A, Lavinha J, Martínez-Jarreta B, Quintana-Murci L, Picornell A, Ramon M, Skorecki K, Behar DM, Calafell F, Jobling MA (December 2008). "The genetic legacy of religious diversity and intolerance: paternal lineages of Christians, Jews, and Muslims in the Iberian Peninsula". American Journal of Human Genetics. 83 (6): 725–36. doi:10.1016/j.ajhg.2008.11.007. PMC 2668061. PMID 19061982.
- ^ L177은 나중에 PF3359 및 동등한 SNP를 위해 폐기되었으며, 2009년 패밀리 트리 DNA에서 처음 확인되었다.L177.1 성분은 Y 염색체 위치 23397163에서, L177.2는 25030912에서, L177.3은 25750264에서 발견된다.자세한 내용은 http://ytree.ftdna.com/index.php?name=Draft&parent=20173662 Archived 2010-10-18 at the Wayback Machine 을 참조하십시오.
- ^ "..Project Rosters - Haplogroup G Project". google.com. Archived from the original on 2013-11-27. Retrieved 2011-12-09.
- ^ Hammer MF, et al. (2009). "Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood". Human Genetics. 126 (5): 707–17. doi:10.1007/s00439-009-0727-5. PMC 2771134. PMID 19669163.
- ^ Habe, M; et al. (March 28, 2012). "Afghanistan's Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events". PLOS ONE. 7 (3): e34288. Bibcode:2012PLoSO...734288H. doi:10.1371/journal.pone.0034288. PMC 3314501. PMID 22470552.
- ^ Copyright 2015 ISOGG. "ISOGG 2015 Y-DNA Haplogroup Tree". isogg.org.
- ^ Underhill PA, Passarino G, Lin AA, Shen P, Mirazón Lahr M, Foley RA, Oefner PJ, Cavalli-Sforza LL (2001). "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. 65 (Pt1): 43–62. doi:10.1046/j.1469-1809.2001.6510043.x. PMID 11415522. S2CID 9441236.
- ^ Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–38. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
- ^ http://ymap.ftdna.com/cgi-bin/gbrowse_details/hs_chrY?name=L240;class=Sequence;ref=ChrY;start=3191153;end=3191153;feature_id=40369[영구 데드링크]
- ^ Sims LM, et al. (2009). Batzer MA (ed.). "Improved Resolution Haplogroup G Phylogeny in the Y Chromosome, Revealed by a Set of Newly Characterized SNPs". PLOS ONE. 4 (6): 1–5. Bibcode:2009PLoSO...4.5792S. doi:10.1371/journal.pone.0005792. PMC 2686153. PMID 19495413.
- ^ King TE, Fortes GG, Balaresque P, Thomas MG, Balding D, Maisano Delser P, Neumann R, Parson W, Knapp M, Walsh S, Tonasso L, Holt J, Kayser M, Appleby J, Forster P, Ekserdjian D, Hofreiter M, Schürer K (2014). "Identification of the remains of King Richard III". Nat. Commun. 5: 5631. Bibcode:2014NatCo...5.5631K. doi:10.1038/ncomms6631. PMC 4268703. PMID 25463651.
- ^ "The Daily Beast".
- ^ "Results from the Hamman Family Y-Chromosome DNA Tests".
- ^ "Haplogroup G2a (Y-chromosomal DNA) - Eupedia".
- ^ "G-P303 (Y-DNA)".
- ^ https://haplogroup.info/all-ancient-dna-full.xlsx[베어 URL 스프레드시트 파일]
외부 링크
- 하플로그룹 G 프로젝트 현장
- 내셔널 지오그래픽의 하플로그룹 G 확산
- Genebase의 Happlogroup G 튜토리얼
- Y-DNA 하플로그룹 G와 금년도 ISOGG 하플로트리의 하위 그룹
- 하플로그룹 G를 사용하는 Y-Search 사용자[영구 데드링크]
- 브리튼 제도 DNA 프로젝트
- 가계도 DNA 하플로그룹 G 프로젝트
- 가계도 DNA에 대한 G Arabia 프로젝트
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Từ khóa » G M201
-
Haplogroup G-M201 - Indo
-
Y-DNA Haplogroup G-M201 - Marres
-
Haplogroup G-M201 | Familypedia - Fandom
-
Haplogroup G-M201 - Wikimedia Tiếng Việt
-
Haplogroup G-M201
-
Where Did The Haplogroup G-M201 Originate? - Quora
-
G_Arabia - FamilyTreeDNA
-
About: Haplogroup G-M201 - DBpedia
-
G Tribe - Did G-M201 Really Originate In The Region Of... - Facebook
-
G YTree - YFull
-
Haplogroup G-M201 - YouTube
-
Haplo-G-M201 [Explicit] By Mike Colin On Amazon Music