Bài viết này cần thêm chú thích nguồn gốc để kiểm chứng thông tin. Mời bạn giúp hoàn thiện bài viết này bằng cách bổ sung chú thích tới các nguồn đáng tin cậy. Các nội dung không có nguồn có thể bị nghi ngờ và xóa bỏ. (Tìm hiểu cách thức và thời điểm xóa thông báo này)
Lá cây: nơi thực hiện quá trình quang hợp ở thực vật.
Quang tổng hợp hay gọi tắt là quang hợp là quá trình thu nhận và chuyển hóa năng lượng ánh sáng Mặt trời của thực vật, tảo và một số vi khuẩn để tạo ra hợp chất hữu cơ phục vụ bản thân cũng như làm nguồn thức ăn cho hầu hết các sinh vật trên Trái Đất. Quang hợp trong thực vật thường liên quan đến chất tố diệp lục màu xanh lá cây và tạo ra oxy như một sản phẩm phụ.[1]
Năng lượng hóa học này được lưu trữ trong các phân tử carbohydrate như đường, và được tổng hợp từ carbon dioxide và nước. Trong hầu hết các trường hợp, oxy cũng được tạo ra như là một sản phẩm phụ. Hầu hết các thực vật, tảo và vi khuẩn cyanobacteria thực hiện quang hợp, và các sinh vật như vậy được gọi là sinh vật quang dưỡng. Quang hợp giúp duy trì nồng độ oxy trong không khí và cung cấp tất cả các hợp chất hữu cơ và hầu hết các năng lượng cần thiết cho sự sống trên Trái Đất.[1]
Mặc dù quá trình quang hợp được thực hiện khác nhau với các loài thực vật khác nhau, quá trình này luôn luôn bắt đầu khi năng lượng từ ánh sáng được hấp thụ bởi các protein được gọi là "trung tâm phản ứng quang hợp" có chứa chất diệp lục (và các Tế bào sắc tố có màu khác). Ở thực vật, các protein này được tổ chức bên trong các bào quan gọi là lục lạp, có nhiều nhất trong các tế bào lá, trong khi ở vi khuẩn các protein này được nhúng vào trong màng sinh chất (màng tế bào). Trong các phản ứng phụ thuộc vào ánh sáng trên, một số năng lượng được sử dụng để tách các electron từ các chất thích hợp như nước, sản xuất khí oxy. Thêm vào đó, hai hợp chất tiếp tục được tạo ra: nicotinamide adenine dinucleotide phosphate (NADPH) và adenosine triphosphate (ATP), các "đơn vị tiền tệ năng lượng" của các tế bào.
Tóm tắt quá trình quang tổng hợp (photosynthesis): quang hợp bao gồm hai pha: pha sáng xảy ra ở thylakoid, còn pha tối xảy ra ở chất nền stroma lục lạp.
Ở thực vật, tảo và vi khuẩn lam, đường được sản xuất bởi một chuỗi các phản ứng không phụ thuộc vào ánh sáng, được gọi là chu trình Calvin, nhưng một số vi khuẩn sử dụng các cơ chế khác nhau, chẳng hạn như chu trình Krebs ngược. Trong chu trình Calvin, khí carbon dioxide được tích hợp vào các hợp chất carbon hữu cơ đã có sẵn, chẳng hạn như ribulose bisphosphate (RuBP).[2] Sử dụng ATP và NADPH được các phản ứng phụ thuộc vào ánh sáng tạo ra, kết quả là các hợp chất này sau đó được giảm và loại bỏ để hình thành carbohydrate cao hơn như glucose.
Trong các chuỗi thức ăn tự nhiên, các sinh vật quang dưỡng (sống nhờ nguồn năng lượng do quang hợp) thường là những mắt xích đầu tiên; nghĩa là các sinh vật còn lại đều sử dụng sản phẩm của quá trình quang hợp phục vụ nhu cầu dinh dưỡng của chúng. Do vậy, quang hợp là chuỗi phản ứng hóa học quan trọng bậc nhất trên Trái Đất, vì nó tạo năng lượng cho sự sống trong sinh quyển. Quá trình quang hợp cũng sản sinh ra khí oxy, tạo nên một bầu khí quyển chứa nhiều oxy cho Trái Đất, một bầu khí quyển vốn dĩ chỉ chứa nitơ và cácbônic trước khi có sinh vật quang dưỡng.
Ở thực vật, quá trình quang hợp chủ yếu được thực hiện nhờ diệp lục (chlorophyll nghĩa là diệp lục; chloro- nghĩa là thứ có màu xanh lục. Sắc tố này thường chứa trong các bào quan gọi là lục lạp. Mặc dù, hầu hết các phần của nhiều loài thực vật đều có màu xanh, năng lượng của quá trình quang hợp chủ yếu được thu nhận từ lá. Quá trình quang hợp của thực vật, tảo và vi khuẩn lam (cyanobacteria) sử dụng chlorophyll và sản sinh ra oxy. Một số loài vi khuẩn quang dưỡng không sử dụng chlorophyll mà dùng một sắc tố tương tự gọi là bacteriochlorophylls và quá trình quang hợp của các vi khuẩn này không sản sinh oxy.
Từ nguyên
[sửa | sửa mã nguồn]
Chữ Hán: 光総合, 光合, tiếng Anh là Photosynthesis bắt nguồn từ tiếng Hy Lạp φῶς: phōs (ánh sáng) và σύνθεσις: synthesis (đặt cùng nhau). Do đó quá trình này có tên quang hợp (光合), gồm hai chữ quang (光) - "ánh sáng", và hợp (合) - "nhóm lại". Tiếng Hy Lạp cũng tương tự, từ φῶς (tức phōs) nghĩa là "ánh sáng", và σύνθεσις (tức synthesis) nghĩa là "tổng hợp lại".[3][4][5]
Lịch sử
[sửa | sửa mã nguồn]
Các sinh vật đầu tiên trên Trái Đất xuất hiện từ cách đây khoảng 3 - 4 tỉ năm tổng hợp thức ăn cho chúng từ những vật chất vô cơ bằng sự hóa tổng hợp (chemosynthesis), tức là lấy năng lượng từ các phản ứng hóa học từ các chất vô cơ như H2, NH4, H2S. Ngày nay, các sinh vật này vẫn còn tồn tại trong những môi trường rất đặc biệt như trong các hố xí, suối nước nóng có lưu huỳnh và các miệng núi lửa trên các sàn đại dương, được gọi là các sinh vật yếm khí. Sau đó xuất hiện nhóm sinh vật có khả năng hấp thu năng lượng ánh sáng mặt trời để tổng hợp ra các hợp chất hữu cơ phức tạp, sự quang tổng hợp (photosynthesis), thường được gọi tắt là sự quang hợp, đây là một quá trình sinh học, chuyển năng lượng ánh sáng thành năng lượng hóa học. Các sinh vật quang hợp đầu tiên này không tạo ra oxy.
Chu trình Calvin
Về sau một số sinh vật có khả năng sử dụng nước cho sự quang hợp, tạo ra O2, dần dần tích tụ trong khí quyển, một số sinh vật tiến hóa khác có khả năng sử dụng O2 xúc tác trong các phản ứng để giải phóng năng lượng trong các phân tử thức ăn. Quá trình này được gọi là sự hô hấp hiếu khí (aerobic respiration). Sự quang hợp sử dụng CO2 và H2O tạo ra từ sự hô hấp hiếu khí và sự hô hấp hiếu khí thì sử dụng thức ăn và O2 sinh ra từ sự quang hợp.
Cả hai loại sinh vật này được gọi chung là sinh vật tự dưỡng-tự tổng hợp chất hữu cơ từ vật chất vô cơ, phân biệt với sinh vật dị dưỡng phải lấy thức ăn hữu cơ từ môi trường chung quanh, chúng tiêu thụ các sinh vật tự dưỡng.
Lục lạp
[sửa | sửa mã nguồn]Cấu tạo của một lục lạp trong tế bào thực vật trong đó (1) granum (2) màng lục lạp bao gồm (3) màng ngoài, (4) xoang gian màng và (5) màng trong. (6) Thylakoid bao gồm (7) xoang thylakoid và (8) màng thylakoid. (10) Chất nền stroma (11) DNA vòng, trần của lục lạp (12) Ribosome (14) Hạt tinh bột
Để hiểu chi tiết về lục lạp, hãy tham khảo bài viết Lục lạp.
Cấu tạo của lục lạp
[sửa | sửa mã nguồn]
Lục lạp là một bào quan đặc biệt của tế bào (đặc biệt là thực vật), giúp chuyển hóa và dự trữ năng lượng photon ánh sáng dưới dạng các liên kết trong phân tử glucose. Giống ty thể, màng lục lạp được cấu tạo bởi hai lớp màng phospholipid kép nhưng màng trong không xẻ khúc mạnh thành các mào, mesosome... mà hai lớp màng khá bằng phẳng. Bên trong lục lạp được bao bọc bởi chất nền stroma, chứa hệ enzyme tham gia vào pha tối quá trình quang hợp.
Thylakoid cấu tạo bởi lớp phospholipid kép, màng thylakoid chứa các phức hệ quang hợp (sắc tố quang hợp), nơi thực hiện chuỗi truyền electron (thẳng hàng hoặc vòng) nhằm bơm proton H+ từ chất nền vào xoang thylakoid để bơm qua protein ATP synthase tổng hợp nên ATP cho nhằm phục vụ cho pha tối quá trình quang hợp. Thylakoid xếp chồng lên nhau tạo thành hạt grana (granum).
Bên cạnh đó, tương tự như ty thể, lục lạp có DNA vòng trần cho nên có khả năng nhân đôi độc lập với nhân tế bào. Ribosome của lục lạp cũng khá đặc biệt là ribosome 70S (giống như ribosome của vi khuẩn) trong đó ribosome của sinh vật nhân thực là 80S.
Ở động vật, do không có lục lạp nên cacbohydrate không được tổng hợp từ lục lạp. Tuy nhiên, ta cũng có bào quan khác tương tự thay thế đó chính là lưới nội chất (ER) trơn, là nơi tổng hợp nên lipid, cacbohydrate cho tế bào, dự trữ cation Ca2+ và khử độc cho tế bào.
Sắc tố quang hợp
[sửa | sửa mã nguồn]
Có khá nhiều loại sắc tố quang hợp như diệp lục, carotenoid, phycoblin, anthocyanine... Hầu hết chúng đều có bản chất là lipid (steroid) nên có tính kị nước do đó chúng hầu như không tan trong nước (ngoại trừ anthocyanine, có trong củ dền, tan mạnh trong nước do nó không có bản chất là lipid). Ở thực vật, sắc tố quang hợp chính là chlorophyll (mà cụ thể là chlorophyll a), các sắc tố phụ như chlorophyll b, caroteinoid, phycobilin... có vai trò hấp thụ năng lượng photon và truyền cho chlorophyll a trung tâm, bên cạnh đó sắc tố phụ cũng góp phần sưởi ấm cho tế bào.
Phổ hấp thụ của sắc tố
[sửa | sửa mã nguồn]
Phổ hấp thụ của sắc tố là câu trả lời thích hợp nhất cho câu hỏi: "Tại sao lá cây có màu xanh?" và đáp án này đến từ chlorophyll của lục lạp trong tế bào lá cây (cụ thể là tế bào mô giậu). Ánh sáng lưỡng tính tức vừa có tính chất sóng vừa có tính chất hạt (hạt photon ánh sáng), và mối quan hệ giữa bước sóng và năng lượng photon tỉ lệ nghịch với nhau. Nghĩa là ánh sáng có bước sóng càng nhỏ, năng lượng photon càng lớn như ánh sáng tím (có bước sóng ngắn trong các vùng ánh sáng nên năng lượng cao gấp đôi ánh sáng đỏ). Ánh sáng trắng là hỗn hợp của nhiều vùng màu xếp từ bước sóng dài đến ngắn là đỏ, cam, vàng, lục, lam, chàm và tím. Chlorophyll hấp thụ ánh sáng đỏ và tím mạnh nhất còn ánh sáng xanh hấp thụ kém nhất (gần như không hấp thụ). Do đó khi chiếu ánh sáng trắng vào chlorophyll thì chỉ có ánh sáng xanh lục không bị hấp thụ và phản xạ lại nên ta nhìn thấy lá cây có màu xanh. Nếu loại bỏ ánh sáng xanh lục thì quá trình quang hợp diễn ra vẫn bình thường, không ảnh hưởng
Pha sáng
[sửa | sửa mã nguồn] Bài chi tiết: Chuỗi chuyền điện tử
Điều kiện xảy ra và bản chất của pha sáng quá trình quang hợp
[sửa | sửa mã nguồn]Ảnh chụp hiển vi điện tử của một lục lạp: chồng grana được tạo bởi thylakoid - nơi xảy ra pha sáng quá trình quang hợp
Pha sáng quá trình quang hợp xảy ra dưới tác dụng của ánh sáng. Ở tế bào thực vật, quá trình này diễn ra ở thylakoid trong lục lạp. Pha sáng thực chất là quá trình phosphoryl hóa (để tổng hợp ATP, adenosine triphosphate) và quá trình tổng hợp nên NADPH nhằm cung cấp năng lượng cho pha tối quá trình quang hợp. Vậy bản chất của pha sáng là chuyển năng lượng từ photon ánh sáng sang phân tử ATP, NADPH2 mà cụ thể là dưới dạng các liên kết hóa học trong ATP (liên kết cao năng của nhóm phosphate) và NADPH2.
Quang hệ PSI và PSII
[sửa | sửa mã nguồn]
Quang hệ là phức hệ của protein với các sắc tố quang hợp. Mỗi quang hệ bao gồm các sắc tố phụ (như carotenoid, chlorophyll b...), đôi chlorophyll a trung tâm và một chất nhận electron sơ cấp. Có hai loại chlorophyll a trung tâm là P680 (tức chlorophyll a hoạt động hiệu quả nhất ở ánh sáng có bước sóng là 680) và P700 (tức chlorophyll a hoạt động tốt nhất ở ánh sáng có bước sóng là 700). Vậy quang hệ PSII thì đôi chlorophyll a trung tâm là P680, còn quang hệ PSI thì đôi chlorophyll a trung tâm là P700 (thứ tự I, II chỉ ra thời điểm phát hiện ra, nhưng quang hệ PSII hoạt động trước quang hệ PSI).
Quang phân ly
[sửa | sửa mã nguồn] Bài chi tiết: Quang phân ly
Xét phương trình đơn giản của quang hợp như sau:
6 CO 2 + 12 H 2 O → c h l o r o p h y l l p h o t o n C 6 H 12 O 6 + 6 O 2 ↑ + 6 H 2 O {\displaystyle {\ce {6CO_2 + 12H_2O ->[{photon}][{chlorophyll}] C_6H_12O_6 + 6O_2 ^ + 6H2O}}}
Trước đây, người ta quan niệm rằng khí oxy sinh ra từ quá trình quang hợp do sự phân hủy của CO2 thành khí O2 và C. Sau đấy C liên kết với các phân tử nước H2O tạo thành đường glucose cần thiết cho sinh vật C6H12O6 và một số carbohydrate Cm(H2O)n như đường fructose C 6 H 12 O 6 {\displaystyle C_{6}H_{12}O_{6}} . Tuy nhiên, một số nhà khoa học đã phản đối quan niệm truyền thống này. Họ tiến hành nhiều thí nghiệm để phản bác lí thuyết ấy và cuối cùng họ đã thành công. Dưới đây là hai thí nghiệm tiêu biểu khẳng định khí O2 không sinh ra từ CO2 mà từ H2O.
Thí nghiệm của van Niel
[sửa | sửa mã nguồn]
Trong những năm 30 của thế kỉ XX, giáo sư C. B van Niel từ trường đại học Stanford, Hoa Kỳ đã tiến hành thí nghiệm như sau: ông nuôi cấy vi khuẩn lưu huỳnh tía Chromatiales trong môi trường có CO2. Vi khuẩn này là vi khuẩn có kiểu dinh dưỡng quang tự dưỡng nhưng không giống thực vật hay vi khuẩn lam, chúng không sử dụng H2O mà thay vào đó là hydro sulfide H2S cho quá trình quang tự dưỡng của chúng như sau:
6 CO 2 + 12 H 2 S → c h l o r o p h y l l p h o t o n C 6 H 12 O 6 + 12 S ↓ + 6 H 2 O {\displaystyle {\ce {6CO2 + 12H2S ->[{photon}][{chlorophyll}] C_6H_12O_6 + 12S v + 6H_2O}}}
Bacteriachlorophyll (khuẩn diệp lục) là một chlorophyll được sử dụng trong quá trình quang hợp ở nhóm vi sinh vật này. Vậy theo quan niệm truyền thống nếu O2 sinh ra từ CO2 thì ở vi khuẩn lưu huỳnh tía người ta phải tìm thấy khí O2 sinh ra. Nhưng ở nhóm vi sinh vật này, CO2 không sinh ra khí O2 mà thay vào đó tạo ra các giọt màu vàng của lưu huỳnh S. Vậy lưu huỳnh được sinh ra do H2S phân li ra S.
Van Niel giải thích rằng vi khuẩn phân ly H2S và dùng các proton H+, electron e- để tạo ra đường và giải phóng O2 như một sản phẩm phụ.
Từ đấy, xét lại phương trình quang hợp của thực vật, tương tự có thể kết luận rằng O2 sinh ra do H2O phân li ra tạo thành hay H2O phân li ra O2 nhằm lấy hydro để tổng hợp nên carbonhydrate và thải khí O2 như sản phẩm phụ. Đồng thời, van Niel cũng rút ra được phương trình chung cho quá trình quang hợp ở sinh vật quang tự dưỡng:
CO 2 + 2 H 2 X ⟶ [ CH 2 O ] + H 2 O + 2 X {\displaystyle {\ce {CO_2 + 2H_2X -> [CH_2O] + H_2O + 2X}}}
Thí nghiệm đánh dấu phóng xạ
[sửa | sửa mã nguồn]
Gần hai mươi năm sau khi van Niel đưa ra giả thuyết, các nhà khoa học đã chứng minh giả thuyết ấy là đúng nhờ sử dụng oxy-18 O 8 18 {\displaystyle {\ce {^{18}_{8}O}}} , một đồng vị nặng để quan sát đường di chuyển của nguyên tử oxy trong quá trình quang hợp. Nếu đánh dấu đồng vị 18O vào nguyên tử oxy trong phân tử CO2 thì khí oxy sinh ra không có 18O mà thay vào đó lại thấy ở carbohydrate C6H12O6 sinh ra và nước H2O giải phóng ra. Nhưng nếu đánh dấu đồng vị 18O vào nguyên tử oxy trong phân tử H2O thì lại chỉ tìm thấy duy nhất 18O khí O2 sinh ra.
Vậy khí O2 sinh ra do H2O phân ly tạo thành.
Phương trình tổng quát của quang phân ly
[sửa | sửa mã nguồn]
H 2 O → q u a n g n ă n g 2 H + + 2 e − + 1 2 O 2 {\displaystyle {\ce {H_2O ->[{quang năng}] 2H^+ + 2e^- + 1/2O_2}}}
Trong đó hai proton H+ tham gia vào chuỗi truyền electron còn 2 electron sẽ bổ sung cho đôi chlorophyll a P680 trung tâm của quang hệ II, còn khí O2 thải ra ngoài môi trường.
Chuỗi truyền electron thẳng hàng
[sửa | sửa mã nguồn]
Chuỗi truyền electron thẳng hàng là chuỗi truyền electron phổ biến ở thực vật. Quá trình ấy được tóm tắt bằng sơ đồ chữ Z như sau:
Photon ánh sáng va chạm vào phân tử sắc tố của quang hệ PSII khiến electron của phân tử ấy bị kích thích bật lên mức năng lượng cao hơn. Nhưng nhanh chóng electron lại bị sụt thế, "rơi" lại vị trí ban đầu và năng lượng tiếp tục chuyền cho phân tử khác. Phân tử khi nhận năng lượng, electron của phân tử ấy bị kích thích và tiếp tục bị bật lên mức năng lượng cao hơn rồi nhanh chóng sụt xuống ban đầu. Quá trình này tiếp diễn nhau tạo thành một chuỗi liên tục đến khi năng lượng được truyền tới cặp phân tử chlorophyll a trung tâm P680 của PSII.
Chuỗi truyền electron thẳng hàng ở màng thylakoids tổng hợp ATP cho pha tối (quang phosphoryl hóa)Đôi phân tử chlorophyll P680 bị kích thích khiến cặp electron bị bật lên mức năng lượng cao hơn. Tuy nhiên, cặp electron này không quay trở về vị trí ban đầu mà bị chất nhận electron sơ cấp hút. Vậy cặp chlorophyll P680 bị mất electron trở thành cation P680+.
Cation P680+ là chất oxy hóa cực mạnh nên H2O phân ly thành hai proton H+, hai electron e- và phân tử O2. Hai electron này được bổ sung cho cation P680+ trở thành phân tử bình thường P680
Cùng lúc ấy, ánh sáng cũng kích thích các phân tử sắc tố của quang hệ PSI đến khi đôi phân tử chlorophyll a trung tâm của PSI là P700 bị kích thích khiến electron bị bật lên chất nhận electron sơ cấp của PSI. Đôi chlorophyll a P700 bị mất electron nên trở thành chất oxy hóa P700+.
Đôi electron từ chất nhận electron sơ cấp của PSII sẽ được chuyền tới đôi P700+ của PSI biến P700+ thành P700 thông qua các chất vận chuyển electron là pheophytin, plastoquinone, phức hợp cytochrome b6f, plastocyanin.
Sự sụt thế của electron thông qua chuỗi chuyền electron cung cấp năng lượng cho sự tổng hợp ATP. Khi electron di chuyển thông qua phức hệ cytochrome đã xác lập sự bơm proton đã xác lập gradient mà về sau được sử dụng cho cơ chế hóa thẩm.
Đôi electron từ chất nhận electron sơ cấp của PSI tiếp tục xuôi theo chuỗi truyền electron thứ hai thông qua protein ferredoxin. (Chuỗi chuyền này không tạo ra sự chênh lệch về nồng độ proton nên không tạo ra ATP).
Enzyme NADP+ reductase xúc tác NADP+ bị khử thành NADPH nhờ đôi electron và hai proton H+.
Pha tối
[sửa | sửa mã nguồn]Khái quát về chu trình Calvin - Benson
Pha tối (Light-independent reaction) của quá trình quang hợp là tập hợp một chuỗi các phản ứng hóa sinh xảy ra ở chất nền (stroma) của lục lạp mà không cần điều kiện ánh sáng (có thể xảy ra trong tối) nhưng lại có quan hệ mật thiết với pha sáng thông qua sản phẩm từ phản ứng sáng là NADPH (Nicotinamide adenine dinucleotide phosphate) và ATP (adenosine triphosphate). Hầu hết phản ứng tối của thực vật gắn liền với chu trình Calvin (gọi đầy đủ là chu trình Calvin - Benson - Bassham hay còn gọi là chu trình C3).
Chu trình Calvin
[sửa | sửa mã nguồn]
Chu trình Calvin được chia làm ba pha cơ bản:
Pha cố định CO2: ba phân tử Ribulose-1,5-diphosphate (viết tắt RuBP) kết hợp với ba phân tử carbon dioxide tạo ra ba phân tử trung gian C6 nhờ enzyme Ribulose-1,5-diphosphate carboxylase oxygenase (viết tắt RuBisCO). Pha cố định CO2Tuy nhiên đường C6 không bền nên dễ dàng phân hủy thành sáu phân tử 3-phosphoglycerate (viết tắt 3-PGA, 3PG, PGA hoặc PGLA). Người ta xem đường 3-PGA là sản phẩm cố định CO2 đầu tiên.
Pha khử: Sáu phân tử 3-PGA, mỗi phân tử nhận thêm một nhóm phosphate từ ATP biến thành 1,3-biphosphoglycerate. Tiếp theo một đôi electron từ NADPH khử 1,3-biophosphateglycerate và mỗi phân tử này mất đi một nhóm phosphate biến thành hai phân tử glyceraldehyde-3-phosphate (viết tắt G3P)Pha khửVậy tạo ra tổng cộng sáu phân tử G3P, một phân tử G3P tách ra khỏi chu trình và tham gia vào tạo glucose và hợp chất hữu cơ khác. Năm phân tử G 3 P {\displaystyle {\ce {G3P}}} còn lại vẫn tiếp tục tham gia chu trình C3.
Pha tái tạo chất nhận CO2 (RuBP) năm phân tử G3P còn lại sẽ biến thành ba phân tử RuBP va tiêu tốn 3 ATP như sau:Pha tái tạo chất nhậnDưới đây trình bày chuỗi phản ứng biến G3P thành RuBP:
Enzyme triose phosphate isomerase sẽ biến tất cả đường G3P ngược lại thành phân tử đường 3-carbon, dihydroxyacetone phosphate (DHAP).
Enzyme adolase và fructose-1,6-bisphosphatase biến đổi một phân tử G3P và một phân tử DHAP thành fructose 6-phosphate (6C, kí hiệu F6P), một nhóm phosphate bị loại bỏ ở phản ứng này.
Cố định phân tử CO2 còn lại, đồng thời sinh ra hai phân tử G3P.
Hai carbon của F6P bị enzyme transketolase loại bỏ, tạo thành erythrose-4-phosphate. Hai phân tử trên transketolase được thêm vào một G3P, tạo thành đường ketose xylulose-5-phosphate (Xu5P)
E4P và một DHAP sẽ được chuyển thành sedoheptulose-1,7-biphosphate (đường C7) thông qua enzyme aldolase.
Enzme Sedoheptulose-1,7-bisphosphatase phân hủy sedoheptulose-1,7-bisphosphate thành sedoheptulose-7-phosphate (viết tắt S7P) đồng thời giải phóng một gốc phosphate vô cơ.
Phân tử CO2 được cố định sinh ra thêm hai phân tử G3P. Đường ketose S7P bị enzyme transketolase loại bỏ 2 carbon tạo ra ribose-5-phosphate (viết tắt R5P) và 2 carbon ấy được chuyển tới một phân tử G3P tạo ra phân tử Xu5P còn lại. Còn lại một phân tử G3P vừa sinh ra ở đầu phản ứng số 7 cùng với ba phân tử pentose được sản sinh sẽ được chuyển hóa thành đường Ru5P.
R5P sẽ chuyển hóa thành ribulose-5-phophate (Ru5P) nhờ enzyme phosphopentose isomerase. Xu5P cũng được chuyển hóa thành Ru5P nhờ enzyme phosphopentose epimerase.
Cuối cùng, enzyme phosphoribulokinase sẽ gắn nhóm phosphate từ ATP vào phân tử RuP tạo thành RuBP.
Vậy thông qua chu trình Calvin, CO2 được sử dụng triệt để nhằm tạo ra sản phẩm cho thực vật đồng thời giải phóng ra ADP (adenosine diphosphate) và NADP+ là nguyên liệu cho pha sáng.
Ngô (Zea mays) là một thực vật C4 tiêu biểu. Trên đây trình bày giải phẫu của lá ngô phù hợp với chức năng sinh lí quang hợp của nó.
Chu trình Hatch-Slack (C4)
[sửa | sửa mã nguồn] Bài chi tiết: Thực vật C4 và Thực vật CAM
Ở một số thực vật như thực vật C4 hay thực vật CAM thì sản phẩm cố định CO2 đầu tiên không phải là 3-PGA mà là một chất khác là hợp chất hữu cơ 4 carbon là oxaloacetate (4C). Chất oxaloacetate tham gia vào một chuỗi phản ứng rồi giải phóng ra CO2. Lúc này CO2 mới tham gia chu trình Calvin. Ở thực vật C4, chu trình Hatch-Slack xảy ra ở tế bào mô giậu còn chu trình calvin xảy ra ở tế bào bao bó mạch. Cụ thể chu trình này như sau:
Phân tử CO2 tác dụng với hợp chất 3 carbon phosphoenol pyruvate (PEP) nhờ enzyme PEP carboxylase tạo ra sản phẩm cố định CO2 là hợp chất 4 carbon oxaloacetate.
Oxaloacetate gắn H+ lấy từ NADPH tạo ra malate (hợp chất 4 carbon)
Malate sẽ bị phân tách thành CO2 và pyruvate (hợp chất 3 carbon). CO2 sẽ đi vào chu trình Calvin.
Pyruvate sẽ tác dụng với ATP tạo ra PEP.Ở thực vật C4, chu trình Hatch-Slack xảy ra ở tế bào mô giậu (tế bào thịt lá) còn chu trình Calvin xảy ra ở tế bào bao bó mạch.Đối thực vật CAM, quá trình pha tối diễn ra ở tế bào chuyên biệt gọi là nhu mô. Cấu tạo của chúng phù hợp với khí hậu nóng bức, khô hạn như hoang mạc, bán hoang mạc, vùng nhiệt đới... tức ban ngày chúng phải đóng khí khổng lại để ngăn chặn mất nước trong cơ thể còn ban đêm thì mới mở khí khổng ra. Tuy nhiên khi đóng khí khổng lại thì chúng không thể hút khí carbonic từ môi trường nên đối với thực vật CAM, chúng sẽ hút khí carbon dioxide vào ban đêm khi khí khổng đang mở. Carbon dioxide sẽ dược dự trữ trong cơ thể dưới dạng malate nhờ chu trình Hatch-Slack, ban ngày thì chúng mới có thể thực hiện chu trình Calvin. Do đó, đối với thực vật CAM: chu trình Hatch-Slack diễn ra vào ban đêm còn chu trình Calvin diễn ra vào ban ngày.
Chu trình Hatch-Slack có tác dụng như dự trữ CO2 trong cơ thể thực vật nhằm cung cấp nguyên liệu CO2 cho chu trình Calvin. Nhờ thế mà thực vật C4 và thực vật CAM sẽ không bị thiếu hụt khí CO2 cho chu trình Calvin. Còn đối với thực vật C3 (tức chất cố định CO2 đầu tiên là 3-PGA) không có chu trình Hatch-Slack có thể thiếu hụt CO2 trong một số trường hợp nhất định và lúc đó sẽ gây ra hiện tượng hô hấp sáng sẽ trình bày rõ ở mục sau.
Chu trình Hatch-Slack được đặt tên nhằm vinh danh hai nhà khoa học là Marshall Davidson Hatch và C. R. Slack, những người đã làm sáng tỏ chúng ở nước Úc vào năm 1966. Chu trình Hatch-Slack cũng thường được gọi phổ biến là chu trình C4.
Hô hấp sáng
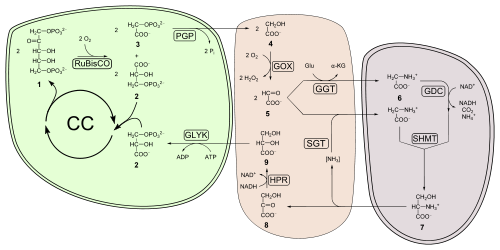
[sửa | sửa mã nguồn] Bài chi tiết: Hô hấp sáng Tóm tắt hô hấp sáng (trong tiếng Anh là Photorespiration) và chu trình Calvin ở thực vật C3.
Trong những ngày khô nóng, thực vật bắt buộc phải đóng khí khổng lại nhằm tránh mất nước hay gây ra hiện tượng xitoriz (hiện tượng xitoriz là hiện tượng xảy ra khi tế bào mất nước quá nhanh do môi trường không khí khô, lúc đó thể tích tế bào giảm nhanh do đó tế bào nhăn nheo lại nhưng chất nguyên sinh vẫn không tách khỏi thành tế bào). Do đó, lá cây không thể hút được khí CO2 từ môi trường bên ngoài. Trong đó chu trình Calvin vẫn tiếp tục sử dụng khí CO2 và quá trình quang phân li trong pha sáng tiếp tục diễn ra.
Vậy, khi đó nồng độ CO2 trong tế bào giảm nhưng nồng độ O2 tiếp tục tăng. Vậy làm thế nào để có thể tăng nồng độ CO2 trong tế bào? Lúc này enzyme RuBisCO sẽ không cố định CO2 vào chất RuBP mà thay vào đó sẽ cố định O2 gây ra hiện tượng hô hấp sáng. Sản phẩm khí của hô hấp sáng bao gồm khí CO2 và NH3.
RuBP tác dụng với khí O2 thông qua enzyme RuBisCO tạo ra 2-phosphoglycolate và 3-PGA (3-PGA sẽ tham gia chu trình Calvin) còn 2-phosphoglycolate sẽ được loại bỏ hai nhóm phosphate vô cơ (Pi) thành glycolate nhờ enzyme phosphoglycolate phosphatase rồi vận chuyển tới bào quan peroxisome trong tế bào.
Glycolate sẽ tác dụng với phân tử O2 dưới tác dụng của enzyme glycolate-oxydase (GOD) sẽ bị chuyển hóa thành glyoxylate đồng thời tạo ra phân tử hydro peroxid H2O2. Phân tử H2O2 nhanh chóng bị enzyme catalase trong peroxisome phân giải thành H2O và O2. Phân tử glyoxylate kết hợp với NH2 sẽ được enzyme glutamate-glyoxylate aminotranferase (GGT) biến đổi thành hai phân tử acid amine glycine.
Phân tử acid amine glycine được vận chuyển tới ty thể, tại đây một phân tử glycine được biến đổi giải phóng CO2, NH4+ đồng thời khử NAD+ thành NADH nhờ enzyme glycine dercarboxylase (GDC) rồi tác dụng với phân tử acid amine glycine còn lại trở thành acid amine serine. Serine được vận chuyển về peroxisome.Chi tiết quá trình hô hấp sáng ở thực vật
Tại peroxisime, acid amine tiếp tục bị enzyme serin-glyoxylate aminotranferase (SGT) biến đổi thành hydroxipyruvate. Hydroxipyruvate tiếp tục bị biến đổi thành glycerate nhờ enzyme hydroxipyruvate reductase (HPR) đồng thời oxy hóa NADH thành NAD+. Glycerate được vận chuyển vào lục lạp trở lại rồi biến đổi thành 3-PGA nhờ enzyme glycerate kinase (GLYK) đồng thời biến đổi ATP thành ADP. Chất 3-PGA tiếp tục tham gia chu trình Calvin.
NH4+ sinh ra từ glycune (mục 3) sẽ vận chuyển về lục lạp rồi kết hợp với 2-oxo-glytarate biến đổi thành acid glutamic, một loại acid amine, nhờ enzyme glutamate synthase - glutamine synthetase. Axit glutamic sẽ bị phân hủy thành NH2 (NH2 tham gia vào quá trình tạo acid amine glycine ở mục 2) và 2-oxo-glytarate. 2-oxo-glytarate sẽ tiếp tục quay trở lại tạo axit glutamic.
Hô hấp sáng làm giảm sản phẩm quang hợp do sử dụng chất nhận CO2 ban đầu RuBP là nguyên liệu cho quá trình oxi hoá trên. Do đó, xét cả ba quá trình trong pha tối, thực vật C4 có năng suất cao nhất còn thực vật CAM có năng suất thấp nhất.
Ý nghĩa và vai trò
[sửa | sửa mã nguồn]
Về mặt năng lượng và dinh dưỡng
[sửa | sửa mã nguồn]
Về mặt năng lượng, quang tổng hợp có bản chất là quá trình hóa vật chất và năng lượng chuyển đổi quang năng thành năng lượng hóa năng và tích trữ trong các liên kết của glucose và các loại đường khác. Do đó về mặt sinh thái, thì mức năng lượng tích trữ trong sinh vật sản xuất (thực vật) là cao nhất. Đồng thời quá trình quang hợp là cửa ngõ để năng lượng được hấp thụ trong hệ sinh thái và di chuyển qua các bậc dinh dưỡng cao hơn.
Về mặt dinh dưỡng - sinh thái, quang hợp là quá trình đồng hóa tổng hợp chất hữu cơ dinh dưỡng từ các chất vô cơ cần thiết cho thực vật, thậm chí còn cung cấp chất dinh dưỡng cho các sinh vật hóa dị dưỡng ăn thực vật. Do đó, thực vật thường là sinh vật sản xuất trong chuỗi và lưới thức ăn. Nếu loại bỏ thực vật ra khỏi chuỗi thức ăn của hệ sinh thái thì có thể khiến cho các sinh vật tiêu thụ khác (trong đó có loài người) không thể tồn tại được.
Về mặt địa hóa - sinh thái, quang hợp là một nhân tố quyết định giúp thực vật có mặt trong chu trình carbon toàn cầu bằng cách hấp thụ carbon dioxide sử dụng trong quá trình tự dưỡng của mình.
Về mặt môi trường
[sửa | sửa mã nguồn]
Khí oxy được thải ra ngoài môi trường thông qua quá trình quang phân li giúp giữ vững nồng độ khí oxy trong khí quyển quanh mức 21%, một lượng đủ và cần thiết để sinh giới tồn tại và phát triển. Đồng thời trong quá trình quang hợp, thực vật còn hút khí CO2 không những tạo ra sản phẩm là tinh bột mà còn giúp điều hòa nồng độ khí CO2 trong khí quyển.
Tham khảo
[sửa | sửa mã nguồn]
^ abBryant DA, Frigaard NU (2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends Microbiol. Quyển 14 số 11. tr. 488–96. doi:10.1016/j.tim.2006.09.001. ISSN 0966-842X. PMID 16997562.
^ Reece J, Urry L, Cain M, Wasserman S, Minorsky P, Jackson R. Biology . Upper Saddle River, NJ: Pearson Education. tr. 235, 244. ISBN 0-321-73975-2. This initial incorporation of carbon into organic compounds is known as carbon fixation.{{Chú thích sách}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
^ "photosynthesis". Online Etymology Dictionary.
^ φῶς. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
^ σύνθεσις. Liddell, Henry George; Scott, Robert; A Greek–English Lexicon at the Perseus Project
Liên kết ngoài
[sửa | sửa mã nguồn] Wikiquote có bộ sưu tập danh ngôn về: Quang hợp
Tư liệu liên quan tới Photosynthesis tại Wikimedia Commons
x
t
s
Thực vật học
Lịch sử thực vật học
Phân ngành
Cổ thực vật học
Cổ di chỉ thực vật học
Thực vật dân tộc học
Hình thái học thực vật
Giải phẫu học thực vật
Phân loại học thực vật
Hệ thống học thực vật
Sinh lý học thực vật
Sinh thái học thực vật
Địa lý thực vật học
Địa thực vật học
Hệ thực vật
Hóa thực vật học
Bệnh học thực vật
Rêu học
Tảo học
Thụ mộc học
Sinh học phát triển tiến hóa thực vật
Thực vật học vũ trụ
Hình thái học thực vật
Tế bào
Vách tế bào
Thể vách
Lạp thể
Cầu sinh chất
Không bào
Mô
Mô phân sinh
Mô dẫn
Bó mạch
Mạch gỗ
Mạch rây
Mô cơ bản
Thịt lá
Tượng tầng
Tầng sinh bần
Tầng sinh mạch
Gỗ
Cơ quan dự trữ
Cấu trúc bề mặt
Bần
Lớp cutin
Lớp sáp
Biểu bì
Khí khổng
Tuyến mật
Hệ thống tiết
Lông, gai
Túm lông
Lông tiết keo
Sinh dưỡng
Rễ
Rễ giả
Rễ biến dạng
Hệ chồi
Chồi
Chồi nách
Chồi bên
Chồi gốc
Chồi nước
Thân
Nhánh
Cuống
Thân hành
Thân rễ
Thân biến dạng
Lá
Cuống lá
Lá kèm
Không cuống
Lá biến dạng
Sinh sản(Bào tử, hoa)
Lá bào tử
Sự phát triển của hoa
Cụm hoa
Cụm hoa vô hạn
Cụm hoa hữu hạn
Lá vảy
Lá bắc
Trục cụm hoa
Cuống hoa
Cán hoa
Hoa
Tiền khai hoa
Vòng
Tính đối xứng của hoa
Hoa đồ
Hoa thức
Đế hoa
Đế hoa rộng
Bao hoa
Tràng hoa
Cánh môi
Đài hoa
Bộ nhụy
Bầu nhụy
Noãn
Đầu nhụy
Túi giao tử cái
Bộ nhị
Nhị
Nhị lép
Bao phấn
Phấn hoa
Khối phấn
Buồng phấn
Trung đới
Hạt phấn
Tầng nuôi dưỡng
Trụ nhị nhụy
Hoa giả
Thể giao tử
Thể bào tử
Phôi
Quả
Giải phẫu quả
Quả đơn
Quả kép
Quả phức
Quả giả
Thai sinh
Bán thai sinh
Hạt
Sự hình thành hạt
Sự phát tán hạt
Dạng sống
Hệ thống Raunkiær
Trạng thái hoa hồng
Thực vật đệm
Dây leo
Thân leo
Cây bụi
Cây bụi lùn
Cây bụi bò lan
Thực vật thân gỗ
Thực vật thân thảo
Thực vật mọng nước
Phân loại học thực vật
Nghiên cứuphân loại họcthực vật
Lịch sử phân loại thực vật
Tập mẫu cây
Phân loại sinh học
Danh pháp thực vật
Tên thực vật
Tên chính xác
Trích dẫn tác giả
Quy tắc Danh pháp của Quốc tế cho tảo, nấm và thực vật (ICN)
- cho Cây Trồng (ICNCP)
Bậc phân loại
Hiệp hội cho Phân loại Thực vật Quốc tế (IAPT)
Hệ thống phân loại thực vật
Hệ thống Takhtajan
Hệ thống APG II
Hệ thống APG III
Phân loại thực vật được gieo trồng
Phân loại cam chanh
Cultigen
Giống cây trồng
Nhóm cây trồng
Giống hoa lan lai
Các nhóm phân loại
Sinh vật lạp thể cổ
Thực vật có phôi
Thực vật không mạch
Rêu
Thực vật có mạch
Dương xỉ & Quyết
Thực vật có hạt
Thực vật hạt trần
Thực vật có hoa
Nhóm phân loại cũ
Tảo
Thực vật tản
Thực vật hoa ẩn
Thực vật thân–rễ
Sinh lý học thực vật
Nguyên liệu
Dinh dưỡng
Quang hợp
Diệp lục
Carotenoid
Carotene
Xanthophyll
Flavonoid
Anthocyanin
Betalain
Hormone thực vật
Ưu thế ngọn
Thoát hơi nước
Áp suất trương
Dòng khối nội bào
Hạt aleurone
Phytomelanin
Đường
Nhựa cây
Tinh bột
Cellulose
Lignin
Tanin
Suberin
Sinh sản
Tiến hóa
Sinh thái học
Xen kẽ thế hệ
Nang bào tử
Bào tử
Nang vi bào tử
Vi bào tử
Nang đại bào tử
Đại bào tử
Thụ phấn
Động vật giao phấn
Ống phấn
Thụ tinh kép
Nảy mầm
Sinh sản sinh dưỡng
Phát triển tiến hóa
Lịch sử tiến hóa
Niên biểu
Từ điển
Thuật ngữ thực vật học
Thuật ngữ hình thái thực vật học
Từ vựng hình thái học thực vật
Thể loại
x
t
s
Sinh thái học: Hệ sinh thái mẫu: Các thành phần dinh dưỡng
Tổng quan
Abiotic component
Abiotic stress
Tập tính
Chu trình sinh địa hóa
Biomass
Biotic component
Biotic stress
Carrying capacity
Competition
Hệ sinh thái
Sinh thái học hệ sinh thái
Mô hình hệ sinh thái
Loài chủ chốt
Tập tính ăn ở động vật
Metabolic theory of ecology
Năng suất
Resource
Sinh vật sản xuất
Hóa tổng hợp
Foundation species
Mixotrophs
Myco-heterotrophy
Mycotroph
Organotrophs
Sinh vật hóa dưỡng
Sinh vật quang dưỡng
Sinh vật quang dị dưỡng
Sinh vật tự dưỡng
Quang hợp
Photosynthetic efficiency
Primary nutritional groups
Primary production
Sinh vật tiêu thụ
Động vật ăn thịt đầu bảng
Bacterivore
Động vật ăn thịt
Chemoorganotroph
Foraging
Generalist and specialist species
Intraguild predation
Động vật ăn cỏ
Sinh vật dị dưỡng
Heterotrophic nutrition
Động vật ăn côn trùng
Mesopredators
Mesopredator release hypothesis
Động vật ăn tạp
Optimal foraging theory
Săn mồi
Prey switching
Sinh vật phân hủy
Chemoorganoheterotrophy
Phân hủy
Sinh vật ăn mùn bã
Detritus
Vi sinh vật
Cổ khuẩn
Thể thực khuẩn
Environmental microbiology
Lithoautotroph
Lithotrophy
Microbial cooperation
Microbial ecology
Microbial food web
Microbial intelligence
Microbial loop
Microbial mat
Microbial metabolism
Phage ecology
Lưới thức ăn
Biomagnification
Ecological efficiency
Kim tự tháp sinh thái
Dòng năng lượng
Chuỗi thức ăn
Bậc dinh dưỡng
Lưới thức ăn điển hình
Lỗ phun lạnh
Miệng phun thủy nhiệt
Intertidal
Rừng tảo bẹ
Hồ
North Pacific Subtropical Gyre
Sông
San Francisco Estuary
Soil
Tide pool
Quá trình
Ascendency
Bioaccumulation
Cascade effect
Climax community
Competitive exclusion principle
Consumer-resource systems
Copiotrophs
Dominance
Ecological network
Diễn thế sinh thái
Chất lượng năng lượng
Energy Systems Language
f-ratio
Hệ số chuyển đổi thức ăn
Feeding frenzy
Mesotrophic soil
Nutrient cycle
Oligotroph
Paradox of the plankton
Trophic cascade
Trophic mutualism
Trophic state index
Phòng ngự/Phản công
Màu sắc động vật
Cơ chế tự vệ của động vật
Ngụy trang
Deimatic behaviour
Herbivore adaptations to plant defense
Bắt chước
Plant defense against herbivory
Predator avoidance in schooling fish
x
t
s
Sinh thái học: Hệ sinh thái mẫu: Các thành phần khác
Sinh thái học quần thể
Abundance
Allee effect
Depensation
Ecological yield
Effective population size
Intraspecific competition
Hàm Lôgit
Mô hình phát triển Malthus
Maximum sustainable yield
Overpopulation in wild animals
Overexploitation
Population cycle
Population dynamics
Population modeling
Population size
Phương trình Lotka–Volterra
Recruitment
Resilience
Small population size
Stability
Các loài
Đa dạng sinh học
Density-dependent inhibition
Ecological effects of biodiversity
Ecological extinction
Các loài đặc hữu
Flagship species
Gradient analysis
Indicator species
Loài du nhập
Loài xâm lấn
Latitudinal gradients in species diversity
Minimum viable population
Neutral theory
Occupancy–abundance relationship
Population viability analysis
Priority effect
Rapoport's rule
Relative abundance distribution
Relative species abundance
Species diversity
Species homogeneity
Species richness
Phân bố loài
Species-area curve
Loài bảo trợ
Tác động giữa các loài
Antibiosis
Tương tác sinh học
Commensalism
Community ecology
Ecological facilitation
Cạnh tranh khác loài
Mutualism
Storage effect
Ký sinh
Cộng sinh
Sinh thái học không gian
Địa lý sinh học
Cross-boundary subsidy
Ecocline
Ecotone
Ecotype
Disturbance
Edge effects
Foster's rule
Habitat fragmentation
Ideal free distribution
Intermediate Disturbance Hypothesis
Island biogeography
Landscape ecology
Landscape epidemiology
Landscape limnology
Metapopulation
Patch dynamics
r/K selection theory
Source–sink dynamics
Ổ
Ổ sinh thái
Ecological trap
Kỹ sư hệ sinh thái
Environmental niche modelling
Guild
Sinh cảnh
Sinh cảnh đại dương
Limiting similarity
Niche apportionment models
Niche construction
Niche differentiation
Các mạng lưới khác
Assembly rules
Bateman's principle
Bioluminescence
Ecological collapse
Ecological debt
Ecological deficit
Ecological energetics
Ecological indicator
Ecological threshold
Ecosystem diversity
Nguyên lý đột sinh
Extinction debt
Kleiber's law
Quy luật cực tiểu của Liebig
Marginal value theorem
Thorson's rule
Xerosere
Khác
Allometry
Alternative stable state
Cân bằng sinh thái
Biological data visualization
Constructal theory
Ecocline
Ecological economics
Dấu chân sinh thái
Ecological forecasting
Ecological humanities
Ecological stoichiometry
Ecopath
Ecosystem based fisheries
Endolith
Evolutionary ecology
Functional ecology
Industrial ecology
Macroecology
Microecosystem
Môi trường tự nhiên
Regime shift
Systems ecology
Urban ecology
Theoretical ecology
x
t
s
Trao đổi chất, dị hóa, đồng hóa
Thông tin chung
Con đường chuyển hóa
Mạng lưới chuyển hóa
Nhóm dinh dưỡng chính
Chuyển hóa năng lượng
Hô hấp tế bào
Đường phân → Khử carbonxyl pyruvate → Chu trình Krebs → Phosphoryl hóa oxy hóa (Chuỗi chuyền điện tử + ATP synthase)
Hô hấp yếm khí
Chất nhận điện tử không phải là oxy
Lên men
Đường phân →
Phosphoryl hóa mức cơ chất
ABE
Ethanol
Acid lactic
Con đường đặc hiệu
Chuyển hóa protein
Tổng hợp protein
Dị hóa
Chuyển hóa carbohydrate(Dị hóa carbohydratevà đồng hóa)





![{\displaystyle {6\,\mathrm {CO} {\vphantom {A}}_{\smash[{t}]{2}}{}+{}12\,\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} {}\mathrel {\xrightarrow[{\text{chlorophyll}}]{\text{photon}}} {}\mathrm {C} {\vphantom {A}}_{\smash[{t}]{6}}\mathrm {H} {\vphantom {A}}_{\smash[{t}]{12}}\mathrm {O} {\vphantom {A}}_{\smash[{t}]{6}}{}+{}6\,\mathrm {O} {\vphantom {A}}_{\smash[{t}]{2}}\uparrow {}{}+{}6\,\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/efb644271d75a3345239572b0f28069d5531e038)

![{\displaystyle {6\,\mathrm {CO} {\vphantom {A}}_{\smash[{t}]{2}}{}+{}12\,\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {S} {}\mathrel {\xrightarrow[{\text{chlorophyll}}]{\text{photon}}} {}\mathrm {C} {\vphantom {A}}_{\smash[{t}]{6}}\mathrm {H} {\vphantom {A}}_{\smash[{t}]{12}}\mathrm {O} {\vphantom {A}}_{\smash[{t}]{6}}{}+{}12\,\mathrm {S} \downarrow {}{}+{}6\,\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/653ede77a06a0b8e1efa062d42e71bd443174877)
![{\displaystyle {\mathrm {CO} {\vphantom {A}}_{\smash[{t}]{2}}{}+{}2\,\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {X} {}\mathrel {\longrightarrow } {}[\mathrm {CH} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} ]{}+{}\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} {}+{}2\,\mathrm {X} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/199c76dffa9a54c54378658bc7fbbd146797c368)
![{\displaystyle {{\vphantom {A}}_{\hphantom {8}}^{\hphantom {18}}{\mkern {-1.5mu}}{\vphantom {A}}_{{\vphantom {2}}{\llap {\smash[{t}]{8}}}}^{{\smash[{t}]{\vphantom {2}}}{\llap {18}}}\mathrm {O} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/736914ec16833d13196f447f0c96e04d45fb69f1)
![{\displaystyle {\mathrm {H} {\vphantom {A}}_{\smash[{t}]{2}}\mathrm {O} {}\mathrel {\xrightarrow {\text{quang năng}} } {}2\,\mathrm {H} {\vphantom {A}}^{+}{}+{}2\,\mathrm {e} {\vphantom {A}}^{-}{}+{}{\mathchoice {\textstyle {\frac {1}{2}}}{\frac {1}{2}}{\frac {1}{2}}{\frac {1}{2}}}\,\mathrm {O} {\vphantom {A}}_{\smash[{t}]{2}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1951fc09fbb7bdef3ff12588b27f77a908f987fe)









![{\displaystyle {\mathrm {G} {\vphantom {A}}_{\smash[{t}]{3}}\mathrm {P} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1d0483ef27d5c9a6f6b33f4bcf55a4ffd86cac20)






 Tư liệu liên quan tới Photosynthesis tại Wikimedia Commons
Tư liệu liên quan tới Photosynthesis tại Wikimedia Commons