Ribosome – Wikipedia Tiếng Việt
Có thể bạn quan tâm
| Sinh học tế bào | |
|---|---|
| Tế bào động vật | |
 Thành phần tế bào động vật điển hình: Thành phần tế bào động vật điển hình:
|
Ribosome là bào quan tổng hợp chuỗi pôlipeptit dựa trên khuôn mã của RNA thông tin.
Đây là một bộ máy phân tử lớn, phức tạp, có mặt trong tất cả các tế bào sống, nơi xảy ra quá trình sinh tổng hợp protein. Ribosome liên kết các amino acid với nhau theo trật tự được quy định bởi phân tử (mRNA). Ribosome bao gồm hai tiểu đơn vị chính - tiểu đơn vị ribosome nhỏ đọc (mRNA), trong khi tiểu đơn vị lớn liên kết các amino acid để tạo thành một chuỗi polypeptide. Mỗi tiểu đơn vị gồm một hoặc nhiều phân tử RNA ribosome (rRNA) và nhiều phân tử protein.
Thuật ngữ "ribosome" có nguồn gốc "ribonucleic acid" (thành phần chính của bào quan) và tiếng Hy Lạp "soma" có nghĩa là "thể".
Trình tự của DNA mã hóa cho một protein có thể được sao chép nhiều lần vào chuỗi RNA thông tin (mRNA) với một trình tự tương tự. Ribosome có thể liên kết với một chuỗi mRNA và sử dụng nó như một khuôn mẫu để xác định chính xác trình tự của các amino acid trong một protein. amino acid được lựa chọn, thu thập và mang đến ribosome bởi RNA vận chuyển (phân tử tRNA), nhập vào một phần của ribosome và liên kết với chuỗi mRNA. Các amino acid sau đó sẽ được liên kết với nhau bởi một phần khác của ribosome. Sau khi protein được sản sinh, nó có thể gấp xoắn lại để tạo nên ra một cấu trúc không gian 3 chiều có chức năng chuyên hóa.
Ribosome được hình thành từ phức hệ RNA và protein nên vì vậy có tên là Ribonucleoprotein. Mỗi ribosome được chia thành hai tiểu đơn vị (hai tiểu phần). Tiểu đơn vị nhỏ hơn liên kết với các khuôn mẫu mRNA, trong khi tiểu đơn vị lớn hơn liên kết với tRNA và các amino acid. Khi ribosome đọc xong một phân tử mRNA, hai tiểu đơn vị này sẽ tách ra. Ribosome còn là một loại enzyme ribozyme, bởi vì có các hoạt động xúc tác phản ứng peptidyl transferase, nhằm mục đích liên kết các amino acid với nhau, thực hiện bởi các RNA ribosome. Ribosome thường được khảm trên những màng nội bào trong hệ thống mạng lưới nội chất hạt.

Ribosome của vi khuẩn, cổ khuẩn và sinh vật nhân thực (ba vực của sự sống trên Trái Đất) có sự khác biệt về kích cỡ, trình tự, cấu trúc và tỷ lệ giữa protein và RNA. Sự sai khác trong cấu trúc cho phép một số thuốc kháng sinh tiêu diệt vi khuẩn bằng cách khống chế ribosome của vi khuẩn mà không ảnh hưởng đến các ribosome của con người. Ở vi khuẩn và vi khuẩn cổ, nhiều ribosome có thể di chuyển dọc theo một chuỗi mRNA cùng một lúc, mà mỗi ribosome "đọc" và tổng hợp một phân tử protein tương ứng. Các ribosome trong ti thể của tế bào nhân thực có chức năng tương tự gần như ribosome vi khuẩn, điều này phản ánh một khả năng về nguồn gốc tiến hóa của ti thể.[1][2]
Khám phá
[sửa | sửa mã nguồn]Ribosome lần đầu tiên được nhà tế bào học người Rumani George Emil Palade quan sát vào giữa thập niên 1950 dưới kính hiển vi điện tử mà chúng hiện lên các hạt một cách dày đặc.[3] Tên gọi "ribosome" do nhà khoa học Richard B. Roberts đề xuất ra vào năm 1958.[4]
Vào năm 1974 cùng với Albert Claude và Christian de Duve, George Emil Palade đã được trao giải Nobel trong lĩnh vực Sinh lý học hay Y học vì phát hiện ra ribosome.[5] Giải thưởng Nobel Hóa học năm 2009 được trao cho Venkatraman Ramakrishnan, Thomas A. Steitz và Ada E. Yonath vì đã xác định được cơ cấu chi tiết và cơ chế hoạt động của ribosome.[6]
Mô tả cấu trúc
[sửa | sửa mã nguồn]

Ribosome bao gồm hai thành phần gắn kết với nhau (Hình 2) và cùng làm việc để chuyển hóa mRNA thành một chuỗi polypeptide trong quá trình tổng hợp protein (Hình 1). Bởi vì chúng được hình thành từ hai thành phần kích thước không bằng nhau, chúng dài một chút trên trục hơn so với đường kính. Ribosome của sinh vật nhân sơ có đường kính khoảng 20 nm (200 Å) và được tạo thành từ 65% RNA ribosome và 35% protein ribosome. Ribosome của sinh vật nhân thực có đường kính từ 25 đến 30 nm (250-300 Å) và tỷ lệ rRNA so với protein là gần như bằng 1. Ribosome trong vi khuẩn bao gồm một hoặc hai chuỗi rRNA. Ribosome của sinh vật nhân thực bao gồm một hoặc ba phân tử rRNA rất lớn (được gọi là ribosome RNA) và nhiều phân tử protein nhỏ hơn. Tinh thể học đã chỉ ra rằng không có protein ribosome gần các mặt phản ứng trong quá trình tổng hợp polypeptide. Điều này cho thấy rằng các thành phần protein của ribosome hoạt động như một lớp tạm thời, có thể tăng cường khả năng của rRNA để tổng hợp protein thay vì trực tiếp tham gia xúc tác (Xem: Ribozyme).
Ribosome chuyển hóa chuỗi polypeptide (nghĩa là protein) từ các cấu trúc di truyền trong phạm vi RNA, bằng cách sử dụng các amino acid cung cấp bởi các RNA vận chuyển (tRNA). Ribosome thoát ra sẽ đính với cytosol (phần bán chất lỏng của tế bào chất), những chất khác bị ràng buộc trong lưới endoplasmic reticulum, xuất hiện của độ nhám; do đó tên của nó còn được gọi là lớp bọc hạt nhân. Mặc dù xúc tác của các liên kết peptide liên quan đến C2 hydroxyl của RNA's P-site (xem phần Function sau đây) adenosine trong một cơ chế trao đổi protein; còn các bước khác trong tổng hợp protein (giống như chuyển vị) được gây ra bởi những thay đổi trong quá trình tái tạo protein. Vì cốt lõi của xúc tác được thực hiện bằng RNA, nên ribosome được phân loại là "ribozymes", và người ta cho rằng nó có thể là phần sót lại cuối cùng của họ RNA.
Ribosome đôi khi được gọi là bào quan, nhưng việc sử dụng từ này thường giới hạn cho việc mô tả tiểu tế bào thành phần bao gồm một màng phospholipid; trong khi ribosome gồm toàn hạt, thì không phải. Vì lý do này, ribosome đôi khi có thể được mô tả như là "bào quan không màng".
Cấu trúc và chức năng của ribosome và những phân tử liên quan, được biết đến như là translational apparatus, đã được quan tâm nghiên cứu từ giữa thế kỷ XX và vẫn là một ngành nghiên cứu rất phổ biến cho đến ngày nay.
Cấu trúc
[sửa | sửa mã nguồn]
Các tiểu đơn vị ribosome của sinh vật nhân sơ và sinh vật nhân thực là khá giống nhau.[8]
Người ta dùng đơn vị đo là Svedberg, một thước đo tốc độ lắng đọng ly tâm chứ không phải là kích thước, và điều này lý giải vì sao các phần tên dù mang số nhưng không có ý nghĩa toán học (ví dụ như 70S được cấu thành bởi 50S và 30S).
Sinh vật nhân sơ có 70S ribosome, mỗi ribosome bao gồm của một tiểu đơn vị nhỏ (30S) và một tiểu đơn vị lớn (50S). Tiểu đơn vị nhỏ này lại có một tiểu đơn vị nhỏ RNA 16S (bao gồm 1540 nucleotide) dính với 21 protein. Tiểu đơn vị lớn bao gồm một tiểu đơn vị RNA 5S (120 nucleotide), một tiểu đơn vị RNA 23S (2900 nucleotide) và 31 protein.[8] Việc đặt tên liên tiếp cho các phần tRNA kết nối trên ribosome E. coli cho phép xác định các protein trên phần A và P có nhiều khả năng liên quan với các hoạt động peptidyl transferase (hình thành liên kết peptid); các protein được đặt tên như vậy là L27, L14, L15, L16, L2, ít nhất là L27 nằm tại bên cho, như E. Collatz và AP Czernilofsky đã cho thấy.[9][10] Các nghiên cứu thêm đã cho thấy rằng các protetin S1 và S21, kết hợp với đuôi 3' của 16S ribosome RNA, có tham gia vào khởi động việc dịch mã.[11]
Sinh vật nhân thực có 80S ribosome, mỗi ribosome bao gồm một tiểu đơn vị nhỏ (40S) và một tiểu đơn vị lớn (60S). Tiểu đơn vị 40S có một RNA 18S (1900 nucleotide) và 33 protein.[12][13] Tiểu đơn vị lớn gồm có một tiểu đơn vị RNA 5S (120 nucleotide), một RNA 28S (4700 nucleotide), và một RNA 5.8S (160 nucleotide) và 46 protein.[8][12][14] Năm 1977, Czernilofsky công bố một nghiên cứu trong đó có sử dụng việc đặt tên liên tiếp để xác định các phần liên kết tRNA trên ribosome gan chuột. Một số protein, bao gồm L32/33, L36, L21, L23, L28/29 và L13 được cho là ở tại hoặc gần trung tâm peptidyl transferase (hình thành liên kết peptid).[15]
Các ribosome được tìm thấy trong lục lạp và ty thể của sinh vật nhân thực cũng bao gồm các tiểu đơn vị lớn và nhỏ liên kết với các protein tạo thành một hạt 70S.[8] Bào quan này được cho là hậu duệ của vi khuẩn (xem thuyết nội cộng sinh) vì các ribosome của nó cũng tương tự như của vi khuẩn.[8]
Các ribosome khác nhau lại có cùng một cấu trúc cốt lõi khá tương đồng mặc dù có kích thước khác biệt rất nhiều. Phần lớn RNA được tổ chức chi tiết vào các chuỗi với cấu trúc bậc ba khác nhau, ví dụ các pseudoknots (nút giả) thể hiện tính đồng trục xếp. RNA thêm trong các ribosome lớn hơn là ở trong các phần chèn thêm dài liên tục, nhờ đó mà chúng tạo thành các vòng từ cấu trúc cốt lõi mà không làm gián đoạn hoặc thay đổi nó. Tất cả các hoạt động xúc tác của ribosome được thực hiện bởi RNA, các protein thì cư trú trên bề mặt và có vẻ có chức năng ổn định cấu trúc.[8]
Sự khác biệt giữa các ribosome vi khuẩn và nhân thực được các nhà hóa học dược phẩm khai thác để tạo ra các loại thuốc kháng sinh có thể tiêu diệt nhiễm trùng do vi khuẩn mà không làm hại các tế bào của người bệnh. Do sự khác biệt trong cấu trúc, các ribosome 70S của vi khuẩn dễ bị kháng sinh tác động trong khi ribosome 80S nhân thực thì không bị ảnh hưởng.[16] Mặc dù ty thể có ribosome tương tự như những vi khuẩn, ti thể không bị ảnh hưởng bởi các loại thuốc kháng sinh bởi vì chúng được bao quanh bảo vệ bởi hai lớp màng ngăn chặn không để kháng sinh vào sâu trong bào quan.[17]
Cấu trúc phân giải cao
[sửa | sửa mã nguồn]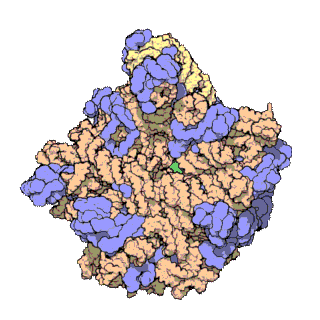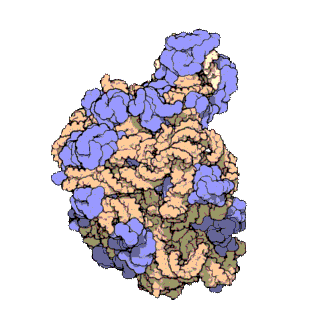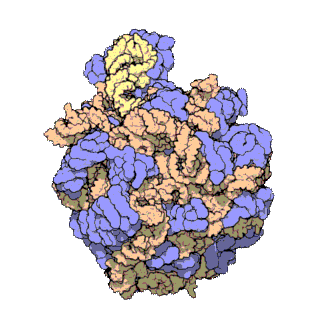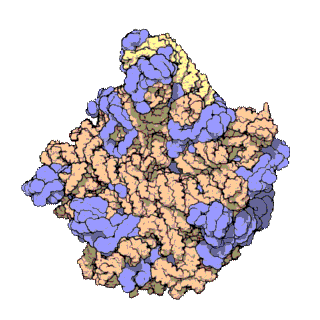
Cấu trúc phân tử chung của ribosome đã được biết đến từ đầu những năm 1970. Đến đầu những năm 2000 cấu trúc đã đạt được xác định ở độ phân giải cao, đến mức một vài Å.
Các nghiên cứu đầu tiên đưa ra cấu trúc của ribosome ở độ phân giải tầm nguyên tử đã được công bố gần như đồng thời vào cuối năm 2000. Các tiểu đơn vị 50S (sinh vật nhân sơ lớn) được xác định từ Haloarcula archaeons marismortui[18] và Deinococcus radiodurans,[19] và cấu trúc của tiểu đơn vị 30S đã được xác định từ Thermus thermophilus.[7] Những nghiên cứu cấu trúc này đã được trao giải Nobel Hóa học năm 2009. Đầu năm sau đó (tháng năm 2001) các nghiên cứu này đã được sử dụng để tái tạo lại toàn bộ hạt T. thermophilus 70S ở độ phân giải 5,5 Å.[20]
Hai bài báo đã được công bố trong tháng 11 năm 2005 với cấu trúc của 70S ribosome của Escherichia coli. Các cấu trúc của một ribosome khuyết đã được xác định ở độ phân giải 3,5-Å bằng cách sử dụng chụp X-quang tinh thể.[21] Hai tuần sau đó, một cấu trúc dựa trên phương pháp hiển vi cryo-electron được công bố,[22] mô tả ribosome tại độ phân giải 11 Å đến 15 Å trong quá trình đưa một sợi protein mới được tổng hợp vào các kênh tạo protein.
Các cấu trúc nguyên tử đầu tiên của ribosome chi tiết hóa với tRNA và phân tử mRNA đã được giải quyết bằng cách sử dụng tinh thể học tia X bởi hai nhóm độc lập, ở 2,8 Å[23] và 3,7 Å.[24] Những cấu trúc này cho phép ta xem chi tiết các tương tác của Thermus thermophilus ribosome với mRNA và tRNA dính tại ở các ribosome điển hình. Tương tác của ribosome với mRNA dài có chứa các chuỗi Shine-Dalgarno sau đó cũng được tạo hình ở độ phân giải 4,5 đến 5,5-Å.[25]
Năm 2011, cấu trúc nguyên tử hoàn chỉnh đầu tiên của ribosome nhân thực 80S từ nấm men Saccharomyces cerevisiae đã thu được bằng phương pháp tinh thể học.[12] Mô hình này cho thấy kiến trúc của các yếu tố nhân sơ và sự tương tác của chúng với lõi được bảo tồn rộng rãi. Đồng thời, các mô hình hoàn chỉnh của một cấu trúc nhân thực ribosome 40S ở Tetrahymena thermophila đã được công bố, mô tả cấu trúc của các tiểu đơn vị 40S cũng như sự tương tác giữa tiểu đơn vị 40S với eIF1 trong thời gian bắt đầu dịch mã.[13] Tương tự như vậy, cấu trúc tiểu đơn vị nhân thực 60S cũng đã được xác định từ Tetrahymena thermophila chi tiết hóa bằng eIF6.[14]
Chức năng
[sửa | sửa mã nguồn]Ribosome là một cấu trúc của tế bào tham gia tổng hợp protein. Protein có vai trò cần thiết cho nhiều chức năng của tế bào như sửa chữa sai hỏng hay điều phối các phản ứng hóa học. Ribosome được tìm thấy bên ngoài nhân tế bào và trôi lơ lửng trong chất tế bào hoặc gắn với mạng lưới nội chất.
Dịch mã
[sửa | sửa mã nguồn] Bài chi tiết: Dịch mã (sinh học)Ribosome là những động cơ chính của quá trình sinh tổng hợp protein, hay còn được biết đến là quá trình dịch mRNA thành protein. mRNA bao gồm một loạt các đơn vị mã truyền cho các ribosome trình tự của các amino acid cần thiết để tạo ra các protein. Sử dụng mRNA như một bản mẫu, ribosome dùng mỗi đơn vị mã (3 nucleotide) của mRNA, ghép nối nó với amino acid thích hợp được cung cấp bởi một aminoacyl-tRNA. aminoacyl-tRNA có chứa một bộ ba đối mã (anti-codon) bổ sung vào một đầu và một amino acid thích hợp ở đầu kia.[26] Các tiểu đơn vị ribosome nhỏ, thường dính vào một aminoacyl-tRNA có chứa các amino acid methionine, liên kết với một AUG đơn vị mã trên mRNA tạo hành các tiểu đơn vị ribosome lớn. Ribosome khi đó có ba phần RNA liên kết, được gọi là A, P và E. Phần A liên kết với một aminoacyl tRNA, phần P liên kết với một peptidyl-tRNA (tRNA dính với peptide được tổng hợp), và phần E liên kết với tRNA tự do trước khi nó ra khỏi ribosome. Tổng hợp protein bắt đầu tại một đơn vị bộ ba mã khởi đầu AUD gần đầu 5' của mRNA. mRNA dính với phần P của ribosome trước. Ribosome có thể để xác định các đơn vị mã khởi đầu bằng cách sử dụng chuỗi Shine-Dalgarno của mRNA nhân sơ và hộp Kozak ở sinh vật nhân thực.
Mặc dù xúc tác của liên kết peptide bao gồm hydroxyl C2 của adenosine P-site của RNA trong cơ chế vận chuyển proton, các bước khác trong quá trình tổng hợp protein (như dịch vị trí) được gây bởi sự thay đổi trong hình thể của protein (conformation). Vì các lõi xúc tác của nó được thực hiện bởi RNA, ribosome được phân loại thành các "ribozyme,"[27] và người ta cho rằng chúng có thể là thành phần còn sót lại của thế giới RNA.[28]

Trong Hình 5, cả các tiểu đơn vị (nhỏ và lớn) của ribosome bắt đầu lắp ráp protein tại bộ ba mã khởi đầu (đầu 5' của RNA). Ribosome sử dụng RNA mà khớp với bộ ba hiện tại trên mRNA để ghép một amino acid vào chuỗi polypeptide. Quá trình được thực hiện đối với mỗi bộ ba trên RNA, trong khi ribosome di chuyển về phía đầu 3' của mRNA. Thông thường trong tế bào vi khuẩn, một vài ribosome làm việc song song với cùng một RNA, quá trình phiên mã và dịch mã diễn ra đồng thời, gọi là polyribosome hay polysome.
Vị trí của Ribosome
[sửa | sửa mã nguồn]Ribosome được phân làm hai loại: "tự do" hoặc "có màng giới hạn".
Ribosome tự do và Ribosome có màng giới hạn chỉ khác nhau trong phân bố không gian của nó và giống hệt nhau về cấu trúc. Cho dù ribosome tồn tại trong trạng thái tự do hoặc có màng giới hạn thì đều phụ thuộc vào sự hiện diện của mạng lưới nội chất (ER) và nhắm đến mục tiêu là những tín hiệu trên protein sẽ được tổng hợp để ribosome có thể là loại có màng giới hạn khi nó đang tạo ra một protein, nhưng cũng có thể là ribosome tự do trong tế bào chất khi nó đang tạo ra một protein khác.
Ribosome đôi khi được gọi là những bào quan, nhưng việc sử dụng thuật ngữ bào quan thường để giới hạn sự miêu tả các thành phần con của tế bào mà bao gồm màng phospholipid, mà ribosome hầu như không có. Vì lý do này mà ribosome đôi khi được miêu tả là các "bào quan không màng".
Ribosome tự do
[sửa | sửa mã nguồn]Loại Ribosome tự do có thể di chuyển bất cứ nơi nào trong bào tương, nhưng không thể di chuyển trong nhân bào và các tế bào quan khác. Protein được hình thành từ các ribosome tự do được phóng vào bào tương và được sử dụng bên trong tế bào. Vì tế bào chất có chứa nồng độ glutathione cao nên trong môi trường khử, các protein có chứa liên kết chất disulfide (được hình thành từ dư lượng chất amino a-xít chứa lưu huỳnh đã bị oxy hóa) không thể được sản sinh ra trong khu vực này.
Ribosome có màng giới hạn
[sửa | sửa mã nguồn]Khi một ribosome bắt đầu tổng hợp các protein cần thiết trong một số cơ quan tế bào, ribosome tạo ra protein này có thể trở thành loại có "màng giới hạn". Trong tế bào nhân thực, việc này sẽ diễn ra trong một khu vực của mạng lưới nội chất (ER) được gọi là "ER thô". Chuỗi polypeptide mới được sản xuất ra sẽ được chèn trực tiếp vào ER bởi sự tổng hợp các vecto thuộc ribosome và sau đó được vận chuyển đến các điểm thông qua con đường bài tiết. Ribosome bị ràng buộc thường sản xuất ra các protein được sử dụng trong màng huyết tương hoặc bị đào thải khỏi tế bào thông qua xuất bào.[29]
Thuyết phát sinh sinh vật
[sửa | sửa mã nguồn] Bài chi tiết: Sinh tổng hợp proteinBên trong tế bào vi khuẩn, ribosome được tổng hợp trong tế bào chất thông qua quá trình phiên mã ở các gen ribosome trong operon. Đối với sinh vật nhân thực, quá trình này được diễn ra ở cả tế bào của tế bào chất và cả trong hạch nhân (một vùng trong nhân tế bào). Quá trình tập hợp là sự kết hợp chức năng của hơn 200 protein trong sự tổng hợp và hoạt động của bốn rRNA, cũng như tập hợp của những rRNA với các protein ribosome.
Tham khảo
[sửa | sửa mã nguồn]- ^ Benne R, Sloof P (1987). "Evolution of the mitochondrial protein synthetic machinery". BioSystems. Quyển 21 số 1. tr. 51–68. doi:10.1016/0303-2647(87)90006-2. PMID 2446672.
- ^ "Ribosomes". Bản gốc lưu trữ ngày 20 tháng 3 năm 2009. Truy cập ngày 28 tháng 4 năm 2011. {{Chú thích web}}: Đã bỏ qua tham số không rõ |= (trợ giúp)
- ^ Palade G. E. (tháng 1 năm 1955). "A small particulate component of the cytoplasm". J Biophys Biochem Cytol. Quyển 1 số 1. tr. 59–68. doi:10.1083/jcb.1.1.59. PMC 2223592. PMID 14381428.
- ^ Roberts, R. B., editor. (1958) "Introduction" in Microsomal Particles and Protein Synthesis. New York: Pergamon Press, Inc.
- ^ "The Nobel Prize in Physiology or Medicine 1974". Nobelprize.org. The Nobel Foundation. Truy cập ngày 10 tháng 12 năm 2012.
- ^ "2009 Nobel Prize in Chemistry". The Nobel Foundation. Truy cập ngày 10 tháng 12 năm 2012.
- ^ a b "Structure of the 30S ribosomal subunit". Nature. Quyển 407 số 6802. tháng 9 năm 2000. tr. 327–39. doi:10.1038/35030006. PMID 11014182. {{Chú thích tạp chí}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ a b c d e f The Molecular Biology of the Cell, fourth edition. Bruce Alberts Garland Science (2002) pg. 342 ISBN 0-8153-3218-1
- ^ Czernilofsky, A; Küchler, E; Stöffler, G.; Czernilofsky, P. (1976). "Site of Reaction on Ribosomal-Protein L27 with an affinity label derivative of transfer-RNA-F(MET)". FEBS Letters. Quyển 63 số 2. Elsevier Science BV. tr. 283–286. doi:10.1016/0014-5793(76)80112-3. PMID 770196.
- ^ Czernilofsky, A; Collatz, E; Stöffler, G; Küchler, E (1974). "Proteins at Transfer-RNA binding-sites of Escherichia-Coli ribosomes". Proceedings of the National Academy of Sciences of the United States of America. Quyển 71 số 1. NAS. tr. 230–234. doi:10.1073/pnas.71.1.230. PMC 387971. PMID 4589893.
- ^ Czernilofsky, A; Kurland, C.G.; Stöffler, G. (1975). "30S ribosomal-proteins associated with 3'-terminus of 16S RNA". FEBS Letters. Quyển 58 số 1. Elsevier Science BV. tr. 281–284. doi:10.1016/0014-5793(75)80279-1. PMID 1225593.
- ^ a b c Ben-Shem A, Garreau de Loubresse N, Melnikov S, Jenner L, Yusupova G, Yusupov M (tháng 2 năm 2011). "The structure of the eukaryotic ribosome at 3.0 Å resolution". Science. Quyển 334 số 6062. tr. 1524–1529. doi:10.1126/science.1212642. PMID 22096102.
- ^ a b Rabl, Leibundgut, Ataide, Haag, Ban (tháng 2 năm 2010). "Crystal Structure of the Eukaryotic 40S Ribosomal Subunit in Complex with Initiation Factor 1". Science. Quyển 331 số 6018. tr. 730–736. doi:10.1126/science.1198308. PMID 21205638.{{Chú thích tạp chí}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b Klinge, Voigts-Hoffmann, Leibundgut, Arpagaus, Ban (tháng 11 năm 2011). "Crystal Structure of the Eukaryotic 60S Ribosomal Subunit in Complex with Initiation Factor 6". Science. Quyển 334 số 6058. tr. 941–948. doi:10.1126/science.1211204. PMID 22052974.{{Chú thích tạp chí}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Czernilofsky, A; Collatz, Ekkehard; Gressner, Axel M.; Wool, Ira G.; Küchler, Ernst (1977). "Identification of tRNA-binding sites on rat-liver ribosomes by affinity labeling". Molecular and General Genetics. Quyển 153 số 3. Springer Verlag. tr. 231–235. doi:10.1007/BF00431588. Truy cập tháng 1 năm 2012. {{Chú thích tạp chí}}: Kiểm tra giá trị ngày tháng trong: |access-date= (trợ giúp)[liên kết hỏng]
- ^ Recht MI, Douthwaite S, Puglisi JD (1999). "Basis for bacterial specificity of action of aminoglycoside antibiotics". EMBO J. Quyển 18 số 11. tr. 3133–8. doi:10.1093/emboj/18.11.3133. PMC 1171394. PMID 10357824.
- ^ O'Brien, T.W. (1971). "The General Occurrence of 55S Ribosomes in Mammalian Liver Mitochondria". J. Biol. Chem. Quyển 245. tr. 3409.
- ^ a b Ban N, Nissen P, Hansen J, Moore P, Steitz T (2000). "The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution". Science. Quyển 289 số 5481. tr. 905–20. doi:10.1126/science.289.5481.905. PMID 10937989.
- ^ Schluenzen F, Tocilj A, Zarivach R, Harms J, Gluehmann M, Janell D, Bashan A, Bartels H, Agmon I, Franceschi F, Yonath A (2000). "Structure of functionally activated small ribosomal subunit at 3.3 Å resolution". Cell. Quyển 102 số 5. tr. 615–23. doi:10.1016/S0092-8674(00)00084-2. PMID 11007480.
- ^ Yusupov MM, Yusupova GZ, Baucom A (tháng 5 năm 2001). "Crystal structure of the ribosome at 5.5 A resolution". Science. Quyển 292 số 5518. tr. 883–96. doi:10.1126/science.1060089. PMID 11283358.
- ^ "Structures of the bacterial ribosome at 3.5 A resolution". Science. Quyển 310 số 5749. tháng 11 năm 2005. tr. 827–34. doi:10.1126/science.1117230. PMID 16272117. {{Chú thích tạp chí}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ "Structure of the E. coli protein-conducting channel bound to a translating ribosome". Nature. Quyển 438 số 7066. tháng 11 năm 2005. tr. 318–24. doi:10.1038/nature04133. PMC 1351281. PMID 16292303. {{Chú thích tạp chí}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ "Structure of the 70S ribosome complexed with mRNA and tRNA". Science. Quyển 313 số 5795. tháng 9 năm 2006. tr. 1935–42. doi:10.1126/science.1131127. PMID 16959973. {{Chú thích tạp chí}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ Korostelev A, Trakhanov S, Laurberg M, Noller HF (tháng 9 năm 2006). "Crystal structure of a 70S ribosome-tRNA complex reveals functional interactions and rearrangements". Cell. Quyển 126 số 6. tr. 1065–77. doi:10.1016/j.cell.2006.08.032. PMID 16962654.
- ^ "Structural basis for messenger RNA movement on the ribosome". Nature. Quyển 444 số 7117. tháng 11 năm 2006. tr. 391–4. doi:10.1038/nature05281. PMID 17051149. {{Chú thích tạp chí}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ Savir, Y; Tlusty, T (ngày 11 tháng 4 năm 2013). "The ribosome as an optimal decoder: a lesson in molecular recognition". Cell. Quyển 153 số 2. tr. 471–9. doi:10.1016/j.cell.2013.03.032. PMID 23582332.
- ^ Rodnina MV, Beringer M, Wintermeyer W (2007). "How ribosomes make peptide bonds". Trends Biochem. Sci. Quyển 32 số 1. tr. 20–6. doi:10.1016/j.tibs.2006.11.007. PMID 17157507.
- ^ Cech T (2000). "Structural biology. The ribosome is a ribozyme". Science. Quyển 289 số 5481. tr. 878–9. doi:10.1126/science.289.5481.878. PMID 10960319.
- ^ Albertyear=2002. The Molecular Biology of the Cell. Garland Science. ISBN 0-8153-4072-9.{{Chú thích sách}}: Quản lý CS1: tên số: danh sách tác giả (liên kết)
Liên kết ngoài
[sửa | sửa mã nguồn]- Lab computer simulates ribosome in motion
- Role of the Ribosome, Gwen V. Childs, copied here
- Ribosome in Proteopedia—The free, collaborative 3D encyclopedia of proteins & other molecules
- Ribosomal proteins families in ExPASy Lưu trữ ngày 30 tháng 4 năm 2011 tại Wayback Machine
- Molecule of the Month Lưu trữ ngày 27 tháng 10 năm 2009 tại Wayback Machine © RCSB Protein Data Bank:
- Ribosome Lưu trữ ngày 14 tháng 11 năm 2010 tại Wayback Machine
- Elongation Factors Lưu trữ ngày 16 tháng 3 năm 2011 tại Wayback Machine
- Palade
- 3D electron microscopy structures of ribosomes at the EM Data Bank (EMDB)
 Bài viết này có sử dụng các nguồn liệu từ Science Primer phát hành tại NCBI, vì đây là một tác phẩm của chính quyền liên bang Hoa Kỳ nên nó thuộc phạm vi công cộng [1].
Bài viết này có sử dụng các nguồn liệu từ Science Primer phát hành tại NCBI, vì đây là một tác phẩm của chính quyền liên bang Hoa Kỳ nên nó thuộc phạm vi công cộng [1].
| |
|---|---|
| Hệ thống nội màng |
|
| Khung xương tế bào |
|
| Nội cộng sinh |
|
| Cấu trúc nội bào khác |
|
| Cấu trúc ngoại bào |
|
Từ khóa » Trong Tế Bào Nhân Thực Riboxom Có ở đâu
-
Cấu Tạo Của Tế Bào Nhân Thực
-
Tế Bào Chất – Wikipedia Tiếng Việt
-
So Sánh Tế Bào Nhân Sơ Và Tế Bào Nhân Thực? - Luật Hoàng Phi
-
Tế Bào Nhân Thực Là Gì? Thành Phần Chính Và Các đặc điểm?
-
Nêu Cấu Tạo Và Chức Năng Của Ribôxôm.
-
Riboxom Có Nhiều Nhất ở đâu - Xây Nhà
-
Bài 8. Tế Bào Nhân Thực - Củng Cố Kiến Thức
-
Tế Bào Nhân Thực Là Gì? Cấu Tạo, Cấu Trúc Tế Bào Nhân Thực
-
Trong Tế Bào, Ribôxôm Có Thể Tìm Thấy ở Trạng Thái Nào Sau đây
-
Ribosome: Nó Là Gì, Chức Năng Của Nó, Những Gì Nó Tạo Ra Và Nó ...
-
Bài 5 Trang 39 SGK Sinh Học 10
-
Tế Bào Nhân Thực Là Gì? Có Và Không Có ở đâu, Khác Gì ...
-
Tế Bào Nhân Thực Có ở đâu
-
Sinh Học 10 Bài 8: Tế Bào Nhân Thực - HOC247