RNA – Wikipedia Tiếng Việt
Có thể bạn quan tâm

Ribonucleic acid (ARN hay RNA) là một phân tử polymer cơ bản có nhiều vai trò sinh học trong mã hóa, dịch mã, điều hòa, và biểu hiện của gen. RNA và DNA là các acid nucleic, và, cùng với lipid, protein và carbohydrat, tạo thành bốn loại đại phân tử cơ sở cho mọi dạng sự sống trên Trái Đất. Giống như DNA, RNA tạo thành từ một chuỗi nucleotide, nhưng không giống DNA là thường tìm thấy nó ở dạng tự nhiên là một sợi đơn gập lại vào chính nó, hơn là sợi xoắn kép. Các sinh vật tế bào sử dụng RNA thông tin (mRNA) đề truyền đạt các thông tin di truyền (sử dụng các base nitric guanine, uracil, adenine, và cytosine, ký hiệu tương ứng bằng các chữ cái G, U, A, và C) cho phép tổng hợp trực tiếp lên các protein chuyên biệt. Nhiều virus mã hóa thông tin di truyền của chúng trong bộ gene RNA.
Một số phân tử RNA đóng vai trò hoạt động bên trong tế bào như là những chất xúc tác cho các phản ứng sinh học, kiểm soát biểu hiện gen, hoặc những đáp ứng cảm nhận và liên lạc trong quá trình truyền tín hiệu tế bào. Một trong những quá trình hoạt động chính là sinh tổng hợp protein, một chức năng phổ biến mà các phân tử RNA trực tiếp tham gia tổng hợp protein trên phân tử ribosome. Quá trình này sử dụng các phân tử RNA vận chuyển (tRNA) mang các amino acid đến phức hệ ribosome, nơi các phân tử RNA ribosome (rRNA) thực hiện ghép nối các amino acid với nhau tạo thành chuỗi tiền protein.
So sánh với DNA
[sửa | sửa mã nguồn]

Cấu trúc hóa học của RNA có những điểm giống với DNA, nhưng có ba điểm khác biệt cơ bản:
- Không như sợi xoắn kép DNA, RNA là phân tử sợi đơn[1] trong hầu hết các chức năng sinh học của nó và chứa chuỗi các nucleotide ngắn hơn nhiều.[2] Tuy nhiên, RNA có thể, bằng cách bắt cặp base bổ sung, tạo thành sợi xoắn kép tự gập từ một sơn đơn, như ở trường hợp tRNA.
- Trong khi "bộ khung" đường-phosphate của DNA chứa deoxyribose, thì bộ khung của RNA là phân tử ribose.[3] Đường ribose có một nhóm hydroxyl gắn với mạch vòng pentose ở vị trí 2', trong khi ở phân tử deoxyribose không có. Nhóm hydroxyl trong bộ khung ribose làm cho RNA ít ổn định so với DNA bởi vì chúng dễ bị thủy phân hơn.
- Base bổ sung của adenine trong DNA là thymine, trong khi ở RNA, nó là uracil, mà là một dạng chưa metyl hóa của thymine.[4]
Giống như DNA, hầu hết các hoạt động sinh học của RNA, bao gồm mRNA, tRNA, rRNA, snRNA, và các RNA không mã hóa khác, chứa các trình tự bổ sung cho phép một phần RNA gập lại[5] và bắt cặp với chính nó để tạo thành sợi kép xoắn ốc. Phân tích những RNA này cho thấy chúng có dạng cấu trúc bậc cao. Không giống như DNA, không chứa một sợi xoắn kép quá dài, mà là một hệ bao gồm các sợi xoắn kép ngắn đính cùng các cấu trúc tương tự như ở protein.
Theo dạng cấu trúc này, RNA có thể trở thành các chất xúc tác (giống như enzyme).[6] Ví dụ, khi xác định cấu trúc của ribosome—một phức hợp RNA-protein tham gia xúc tác hình thành chuỗi peptide—các nhà sinh học phát hiện thấy vị trí hoạt động của nó chứa hoàn toàn của RNA.[7]
Cấu trúc
[sửa | sửa mã nguồn] Bài chi tiết: Cấu trúc acid nucleic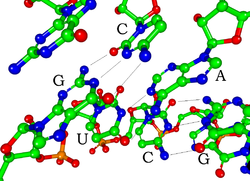
Mỗi nucleotide trong RNA chứa một đường ribose, với carbon được đánh thứ tự từ 1' đến 5'. Nhìn chung, một base được gắn vào vị trí 1' là adenine (A), cytosine (C), guanine (G), hoặc uracil (U). Adenine và guanine là các purine, cytosine và uracil là các pyrimidine. Một nhóm phosphat gắn vào vị trí 3' của một đường ribose và vào vị trí 5' của đường ribose tiếp theo. Nhóm phosphat tích điện âm, khiến cho RNA là phân tử mang điện (polyanion). Các base tạo thành liên kết hiđrô giữa các cytosine và guanine, giữa adenine và uracil và giữa guanine và uracil.[8] Tuy thế, cũng có thể có những tương tác khác, như một nhóm base adenine liên kết với một nhóm khác trong chỗ phình,[9] hoặc tại vòng bốn (tetraloop) GNRA có liên kết cặp base guanine–adenine.[8]

Một thành phần cấu trúc quan trọng của RNA khác biệt với DNA đó là sự có mặt của nhóm hydroxyl tại vị trí 2' trong đường ribose. Sự có mặt của nhóm chức này làm cho dạng xoắn của RNA có dạng A-hình học (A-form geometry),[10] mặc dù trong trường hợp sợi đơn dinucleotide, có thể hiếm gặp RNA trong dạng B-hình học như quan sát thấy ở hầu hết DNA.[11] Dạng A-hình học khiến cho trên phân tử RNA có rãnh (groove) lớn hẹp và rất sâu và một rãnh nhỏ rộng và nông.[12] Hệ quả thứ hai của sự có mặt nhóm 2'-hydroxyl đó là trong các vùng có hình dáng linh hoạt (conformationally flexible regions) của một phân tử RNA (tức là không tham gia vào sự tạo thành sợi xoắn kép), có thể tấn công hóa học vào liên kết phosphodiester bên cạnh để cắt bộ khung RNA.[13]

RNA được phiên mã chỉ ở bốn base (adenine, cytosine, guanine và uracil),[14] nhưng các base này và nhóm đường gắn cùng có thể được chỉnh sửa theo nhiều cách khi RNA trưởng thành. Ở pseudouridine (Ψ), mà trong đó mối liên kết giữa uracil và ribose bị chuyển từ liên kết C–N thành liên kết C–C, và ribothymidine (T) được tìm thấy ở nhiều nơi (nổi bật nhất là nó xuất hiện ở vòng TΨC của tRNA).[15] Một ví dụ base biến đổi khác đó là hypoxanthine, một base adenine đã khử amin mà nucleoside của nó được gọi là inosine (I). Inosine đóng vai trò quan trọng trong giả thuyết cặp base linh hoạt (wobble hypothesis) của mã di truyền.[16]
Có hơn 100 nucleoside biến đổi xuất hiện trong tự nhiên.[17] Sự đa dạng lớn nhất trong cấu trúc của sửa đổi này có thể tìm thấy ở tRNA,[18] trong khi pseudouridine và nucleoside với 2'-O-methylribose thường có mặt trong rRNA là dạng phổ biến nhất.[19] Các nhà sinh học vẫn chưa hiểu đầy đủ vai trò đặc trưng của nhiều biến đổi này trong RNA. Tuy nhiên, đáng chú ý là, trong RNA ribosome, nhiều thay đổi sau phiên mã xảy ra ở những vùng có chức năng cao như trung tâm peptidyl transferase và giao diện tiểu đơn vị, ngụ ý rằng chúng quan trọng đối với chức năng bình thường.[20]
Dạng chức năng của các phân tử RNA sợi đơn, giống như các protein, thường đòi hỏi một cấu trúc bậc ba cụ thể. Các bộ khung cho cấu trúc này được cung cấp bởi các yếu tố cấu trúc bậc hai là liên kết hydro trong phân tử. Điều này dẫn đến một số "miền" có thể nhận biết được của cấu trúc bậc hai như vòng kẹp tóc (hairpin loop), phình và vòng lặp nội bộ (internal loop).[21] Vì RNA mang điện tích, các ion kim loại như Mg2+ cần thiết có mặt để ổn định nhiều cấu trúc bậc hai và bậc ba của RNA.[22]
Dạng đồng phân lập thể enantiomer xuất hiện tự nhiên của RNA là D-RNA chứa các D-ribonucleotide. Mọi trung tâm đối xứng đều nằm trong D-ribose. Bằng cách sử dụng L-ribose hoặc L-ribonucleotide, có thể tổng hợp được L-RNA.L-RNA có tính ổn định lớn hơn chống lại sự thoái biến của RNase.[23]
Giống như các phân tử sinh học có cấu trúc khác như protein, có thể định nghĩa tô pô của một phân tử RNA đã gập. Điều này thường dựa trên sự sắp xếp các vị trí tiếp xúc nội chuỗi bên trong RNA đã gập, gọi là mạch tô pô (circuit topology).
Tổng hợp
[sửa | sửa mã nguồn]Quá trình tổng hợp RNA gọi là phiên mã, luôn cần sự xúc tác của enzym RNA polymerase sử dụng một mạch khuôn của gen trên DNA. Sự khởi đầu phiên mã bắt đầu bằng enzyme gắn kết vào trình tự khởi động trong DNA ở phía "thượng nguồn" của gen.
- Chuỗi xoắn kép DNA ở vùng có gen cần phiên mã đầu tiên phải được tháo xoắn nhờ topoisomerase, sau đó được dãn mạch và tách đôi nhờ enzym helicase.
- Enzym RNA polymerase trượt dọc theo sợi khuôn mẫu (mạch gốc) theo chiều 3’ đến 5’ của gen, tổng hợp lên chuỗi polyribonucleotide theo nguyên tắc bổ sung, được kéo dài theo hướng 5’ đến 3’ (ngược lại với hướng di chuyển của enzym này). Trình tự các deoxyribonucleotide trên mạch gốc của gen không chỉ quyết định trình tự chuỗi polyribonucleotide của RNA, mà còn quy định cả sự kết thúc của quá trình phiên mã.[24]
- Ở tế bào nhân thực, RNA vừa được phiên mã mới chỉ là tiền RNA (pre RNA) hay RNA sơ khai. Nó phải trải qua quá một quá trình gọi là biến đổi sau phiên mã mới tạo nên RNA trưởng thành. Trong quá trình xử lý RNA:
- Đầu 5' của nó được gắn "chóp" GTP, còn đầu 3' của nó sẽ được gắn "đuôi" là pôlyA.
- RNA sơ khai cần phải được cắt bỏ hết các intron (vùng không mã hóa), rồi các intron này sẽ bị phân giải;
- Các intron (đoạn không có mã) của nó bị cắt bỏ, còn các exon (đoạn có mã) sẽ nối với nhau tạo thành một chuỗi bộ ba mã di truyền liên tục. Giai đoạn này gọi là cắt nối RNA.
Quá trình trên được thực hiện nhờ nhiều nhân tố, nhưng quan trọng nhất là spliceosome (thể chế biến) là một tổ hợp phân tử lớn và phức tạp. Sau khi chế biến hoàn tất, RNA trưởng thành được tạo ra và mới được xuất ra tế bào chất qua lỗ nhân.
Ở một số ít nhóm sinh vật, còn có một số RNA polymerase phụ thuộc RNA (RNA-dependent RNA polymerase) sử dụng RNA làm khuôn mẫu cho tổng hợp lên sợi RNA mới. Ví dụ, một số virus RNA (như poliovirus) sử dụng loại enzyme này để sao chép vật liệu di truyền của chúng.[25] Cũng vậy, RNA polymerase phụ thuộc RNA là một phần trong lộ trình can thiệp RNA ở nhiều sinh vật.[26]
Các loại RNA
[sửa | sửa mã nguồn] Xem thêm: Danh sách RNATổng quan
[sửa | sửa mã nguồn]
RNA thông tin (mRNA) là RNA mang thông tin từ DNA đến ribosome, các vị trí dịch mã để sinh tổng hợp protein trong tế bào. Trình tự mã hóa của mRNA xác định lên trình tự amino acid trong protein được tổng hợp ra.[27] Tuy nhiên, nhiều RNA không có vai trò mã hóa cho protein (khoảng 97% sản phẩm RNA từ quá trình phiên mã là những protein không mã hóa trong sinh vật nhân thực[28][29][30][31]).
Những RNA không mã hóa ("ncRNA") này có thể được mã bởi chính bộ gene của chúng (RNA gene), nhưng cũng có thể được tạo thành từ các intron mRNA.[32] Ví dụ nổi bật nhất cho các RNA không mã hóa đó là RNA vận chuyển (tRNA) và RNA ribosome (rRNA), mà cả hai đều tham gia vào quá trình dịch mã.[4] Có các RNA không mã hóa tham gia vào điều hòa biểu hiện gene, xử lý RNA và các vai trò khác. Một số RNA có thểm làm chất xúc tác cho phản ứng sinh hóa như cắt và nối các phân tử RNA khác,[33] và xúc tác tạo thành liên kết peptide trong ribosome;[7] chúng được biết với tên gọi ribozyme.
Theo độ dài
[sửa | sửa mã nguồn]Nếu phân theo độ dài của một chuỗi RNA, có thể chia RNA thành các RNA nhỏ và RNA dài.[34] Bình thường, các RNA nhỏ có độ dài ngắn hơn 200 nt, và các RNA dài có độ dài hơn 200 nt.[35] Các phân tử RNA dài, hay còn gọi là RNA lớn, chủ yếu bao gồm các RNA không mã hóa dài (lncRNA) và mRNA. Phân tử RNA nhỏ bao gồm chủ yếu tiểu đơn vị 5.8S RNA ribosome (rRNA), 5S rRNA, RNA vận chuyển (tRNA), microRNA (miRNA), RNA can thiệp nhỏ (small interfering RNA, siRNA), RNA neucleolar nhỏ (small nucleolar RNA, snoRNAs), RNA tương tác Piwi (Piwi-interacting RNA, piRNA), RNA nhỏ bắt nguồn từ tRNA (tRNA-derived small RNA, tsRNA)[36] và RNA nhỏ bắt nguồn từ rDNA (small rDNA-derived RNA, srRNA).[37]
Theo dịch mã
[sửa | sửa mã nguồn]
RNA thông tin (mRNA) mang các thông tin di truyền về trình tự của một protein đến ribosome, nhà máy tổng hợp protein bên trong tế bào. Nó mã hóa sao cho cứ mỗi ba nucleotide (bộ ba mã hóa hay một codon) tương ứng với một amino acid. Trong tế bào sinh vật nhân thực, một phân tử tiền mRNA (pre-mRNA) được phiên mã từ DNA, sau đó nó được xử lý để trở thành mRNA trưởng thành. Quá trình này bao loại bỏ các đoạn intron—các vùng không mã hóa của pre-mRNA. Sau đó mRNA được đẩy từ nhân tế bào vào bào tương, nơi nó sẽ tìm đến các ribosome và thực hiện dịch mã thành protein tương ứng với sự tham gia cùng tRNA. Trong tế bào sinh vật nhân sơ, mà không có nhân và các gian xoang bào, mRNA có thể liên kết ngay với ribosome trong khi nó đang được phiên mã từ DNA. Sau một thời gian nhất định, các phân tử thông tin này thoái hóa thành các thành phần nucleotide với sự trợ giúp của ribonuclease.[27]
RNA vận chuyển (tRNA) là một sợi RNA nhỏ dài khoảng 80 nucleotide mà vận chuyển một loại amino acid nhất định đến gắn vào chuỗi polypeptide đang dài dần tại vị trí của ribosome đang tổng hợp lên protein trong quá trình dịch mã. Nó có các vị trí cho phép gắn amino acid và một vùng codon đối mã (anticodon) cho phép nhận ra codon gắn trên mRNA thông tin thông qua liên kết hydro.[32]
RNA ribosome (rRNA) là thành phần xúc tác của ribosome. Ribosome ở sinh vật nhân thực chứa bốn loại phân tử rRNA khác nhau: 18S, 5.8S, 28S và 5S rRNA. Ba phân tử rRNA được tổng hợp trong nhân con, và phân tử còn lại được tổng hợp ở nơi khác. Trong bào tương, RNA ribosome và protein kết hợp lại thành phức hệ nucleoprotein gọi là ribosome. Ribosome gắn với mRNA và thực hiện quá trình tổng hợp protein. Một số ribosome thường lúc nào cũng gắn với một sợi mRNA.[27] Gần như mọi RNA tìm thấy trong mọi tế bào sinh vật nhân thực là rRNA.
RNA thông tin-vận chuyển (transfer-messenger RNA, tmRNA) được tìm thấy ở nhiều vi khuẩn và lạp thể. Nó đánh dấu các protein mã hóa bởi mRNAs mà thiếu những codon kết thúc cho sự thoái hóa và ngăn cản ribosome khỏi bị dừng.[38]
RNA điều hòa
[sửa | sửa mã nguồn]Một vài loại RNA có khả năng điều hòa làm sụt giảm quá trình biểu hiện gene bằng cách gắn bổ sung vào một phần của mRNA hoặc đoạn DNA của gene.[39][40] Các microRNA (miRNA; dài 21-22 nt) đã được tìm thấy ở sinh vật nhân thực và tác động thông qua can thiệp RNA (RNAi), nơi một phức hệ bộ phận tác động của miRNA và các enzyme có thể cắt mRNA, cản trở mRNA đang trong quá trình dịch mã, hoặc làm tăng tốc sự thoái hóa của nó.[41][42]
Trong khi các RNA can thiệp nhỏ (small interfering RNA, siRNA; 20-25 nt) thường được tạo ra bằng cách phá vỡ RNA của virus, cũng có những nguồn nội sinh siRNA.[43][44] siRNAs hoạt động thông qua quá trình can thiệp RNA theo cách tương tự như miRNA. Một số miRNAs và siRNAs có thể gây cho các gene chúng tác động tới bị methyl hóa, do đó làm giảm hoặc tăng hoạt động phiên mã ở các gene này.[45][46][47] Ở những động vật có RNA tương tác Piwi (piRNA; 29-30 nt) mà hoạt động trong các tế bào dòng mầm (germline) và được cho là những phân tử phòng thủ chống lại transposon và đóng vai trò quan trọng trong sự hình thành giao tử (gametogenesis).[48][49]
Nhiều sinh vật nhân sơ có các RNA CRISPR, một hệ thống điều hòa tương tự như của can thiệp RNA.[50] Các RNA đối nghĩa (antisense RNA) được lan rộng; mà hầu hết điều hòa làm giảm sự hoạt động của một gene, nhưng có một số là những phân tử kích hoạt quá trình phiên mã.[51] Một cách RNA đối nghĩa hoạt động là khi nó gắn vào một mRNA, tạo thành sợi kép RNA mà chức năng enzyme bị suy giảm đi.[52] Có nhiều RNA không mã hóa sợi dài tham gia điều hòa gene ở sinh vật nhân thực,[53] ví dụ những RNA như thế là Xist, mà nó bao lấy nhiễm sắc thể X ở con cái trong động vật có vú và bất hoạt nó.[54]
Một mRNA có thể chứa những phần tử điều hòa trong chính nó, như các đoạn riboswitch, nằm trong vùng đầu 5' không được dịch mã hoặc vùng đầu 3' không được dịch mã; các yếu tố điều hòa trong vùng (cis-regulatory element) này điều hòa sự hoạt động của chính mRNA.[55] Những vùng không tham gia dịch mã cũng có thể chứa các đoạn mà tham gia vào điều hòa ở các gene khác.[56]
Theo xử lý RNA
[sửa | sửa mã nguồn]
Nhiều RNA tham gia vào sửa đổi các RNA khác. Những đoạn intron bị cắt ra khỏi pre-mRNA bởi spliceosome, mà trong nó chứa một vài RNA hạt nhân nhỏ (small nuclear RNA, snRNA),[4] hoặc các intron có thể là ribozyme mà dùng để cắt chính những đoạn intron khác.[57] RNA cũng có thể được chỉnh sửa bằng dùng các nucleotide A, C, G và U trong một RNA này để thay đổi các nucleotide trong một RNA khác. Ở sinh vật nhân thực, sự chỉnh sửa các nucleotide của RNA nói chung được điều khiển bởi các RNA nucleolar nhỏ (small nucleolar RNA, snoRNA; 60–300 nt),[32] được tìm thấy trong nhân con và các thể Cajal. snoRNAs phối hợp với các enzyme đến một vị trí trên RNA bằng cách bắt cặp base với RNA. Các enzyme này sau đó thực hiện sửa đổi nucleotide. rRNA và tRNA là những phân tử được sửa đổi rất nhiều, nhưng snRNA và mRNA cũng có thể là những mục tiêu cho sửa đổi base.[58][59] RNA cũng có thể bị methyl hóa.[60][61]
Bộ gene RNA
[sửa | sửa mã nguồn]Giống như DNA, RNA có thể được dùng để mang thông tin di truyền. Các virus RNA có bộ gene chứa RNA mã hóa cho các protein của chúng. Bộ gene virus được tái bản bằng một số protein này, trong khi các protein khác có chức năng bảo vệ bộ gene khi hạt virus chuyển sang tế bào vật chủ mới. Viroid là một nhóm thể sinh bệnh khác, nhưng chúng chỉ chứa RNA, và không mã hóa cho bất kỳ một protein nào và được sao chép nhờ các polymerase của tế bào thực vật chủ.[62]
Theo phiên mã ngược
[sửa | sửa mã nguồn]Các virus có bộ gen là RNA phải được tổng hợp ngược trở lại thành DNA nhờ enzyme phiên mã ngược; từ đó tạo nên DNA bổ sung rồi sau đó mới được phiên mã thành những RNA mới để làm khuôn dịch mã. Retrotransposon cũng được lan rộng nhờ cách sao chép DNA và RNA từ tế bào này sang tế bào khác,[63] và telomerase chứa một RNA được sử dụng làm khuôn mẫu cho việc lắp ráp những đoạn cuối của nhiễm sắc thể ở sinh vật nhân thực.[64]
RNA sợi kép
[sửa | sửa mã nguồn]RNA sợi kép (dsRNA) là RNA mà có hai sợi bổ sung, tương tự như ở DNA trong mọi tế bào. dsRNA tạo thành vật liệu di truyền ở một số virus (virus có RNA sợi kép, double-stranded RNA viruses). RNA sợi kép chẳng hạn như ở RNA virus hoặc siRNA có thể kích hoạt can thiệp RNA ở sinh vật nhân thực, cũng như hoạt hóa các protein interferon trong động vật có xương sống.[65][66][67][68]
RNA vòng
[sửa | sửa mã nguồn]Cuối thập niên 1990, các nhà sinh học đã phát hiện có một loại sợi đơn RNA khép kín ở động vật. Sau đó loại này được chính thức xác nhận và gọi là RNA vòng (circRNA). Xem chi tiết về loại này ở trang RNA vòng.
Các khám phá quan trọng về RNA sinh học
[sửa | sửa mã nguồn]
Nghiên cứu về RNA đã dẫn đến nhiều khám phá sinh học quan trọng cũng như nhiều giải Nobel. Acid nucleic được Friedrich Miescher khám phá ra lần đầu tiên vào năm 1868, khi ông gọi các vật liệu này là 'nuclein' do chúng được tìm thấy trong nhân tế bào.[69] Sau đó người ta khám phá ra tại các tế bào sinh vật nhân sơ, mà không có nhân, cũng thấy chứa acid nucleic. Giải thuyết về vai trò của RNA trong sinh tổng hợp protein đã được nêu ra từ năm 1939.[70] Severo Ochoa nhận giải Nobel Sinh lý học và Y khoa năm 1959 (cùng với Arthur Kornberg) cho khám phá của ông về một enzyme cho phép tổng hợp được RNA trong phòng thí nghiệm.[71] Tuy nhiên, loại enzyme khám phá bởi Ochoa (polynucleotide phosphorylase) sau này được chứng minh là có vai trò làm thoái hóa RNA, chứ không phải tổng hợp lên RNA. Năm 1956 Alex Rich và David Davies cho lai hai dòng RNA để tạo thành tinh thể RNA đầu tiên mà cấu trúc của nó có thể xác định bằng kỹ thuật nhiễu xạ tia X (tinh thể học tia X).[72]
Trình tự của 77 nucleotide trong tRNA của một loài nấm men được Robert W. Holley xác định lần đầu tiên vào năm 1965,[73] giúp Holley đoạt giải Nobel Sinh lý học và Y khoa 1968 (cùng với Har Gobind Khorana và Marshall Nirenberg).
Trong đầu thập niên 1970, các retrovirus và enzyme phiên mã ngược được phát hiện, và lần đầu tiên chứng tỏ rằng các enzyme tham gia quá trình sao chép từ RNA vào DNA (quá trình ngược so với chu trình thông thường của sự truyền thông tin di truyền). Nhờ khám phá này, David Baltimore, Renato Dulbecco và Howard Temin được trao giải Nobel Y học năm 1975.
Năm 1976, Walter Fiers cùng các đồng nghiệp lần đầu tiên đã giải trình tự thành công RNA trong một bộ gene của virus, hay bacteriophage MS2.[74]
Năm 1977, các intron và quá trình ghép RNA (RNA splicing) được phát hiện ở cả virus trên động vật và ở gene tế bào, đưa Philip Sharp và Richard Roberts đến giải Nobel năm 1993. Các phân tử RNA xúc tác (ribozyme) được phát hiện vào đầu thập kỷ 1980, và mang lại cho Thomas Cech và Sidney Altman giải Nobel năm 1989. Năm 1990, người ta tìm thấy trong thực vật Petunia (dã yên thảo) là có thể dùng các gene để tắt các gene tương tự trong chính loài thực vật này, một khám phá đã mở đường cho kỹ thuật can thiệp RNA sau này.[75][76]
Trong khoảng cùng thời gian này, các sợi RNA dài 22 nt, mà hiện nay gọi là microRNA, được tìm thấy có vai trò trong sự phát triển của C. elegans.[77]
Nghiên cứu can thiệp RNA đưa đến giải Nobel Y học năm 2006 cho Andrew Fire và Craig Mello, và giải Nobel Hóa học cho nghiên cứu về quá trình phiên mã RNA trao cho Roger Kornberg trong cùng năm. Sự khám phá các RNA điều hòa biểu hiện gene đã dẫn đến những nỗ lực phát triển các loại thuốc là từ RNA, như siRNA, có chức năng làm tắt một số gene.[78]
Liên quan đến hóa học tiền sinh học và thuyết phát sinh sinh vật
[sửa | sửa mã nguồn]Năm 1967, Carl Woese nêu ra giả thuyết rằng RNA có thể là chất xúc tác và gợi ý những dạng sống nguyên thủy nhất (các phân tử tự tái bản) có thể dựa trên RNA cả về mặt chứa đựng thông tin di truyền và làm chất xúc tác cho các phản ứng hóa sinh—hay còn gọi là giả thuyết thế giới RNA.[79][80]
Tháng 3 năm 2015, các nucleotide phức tạp của DNA và RNA, bao gồm uracil, cytosine và thymine, được thông báo là đã tổng hợp được trong phòng thí nghiệm dưới những điều kiện của không gian ngoài thiên thể, sử dụng các hóa chất ban đầu, như pyrimidine, một hợp chất hữu cơ phổ biến tìm thấy trong các vẫn thạch. Pyrimidine, giống như các hydrocarbon thơm đa vòng (polycyclic aromatic hydrocarbons, PAHs), là một trong những hợp chất giàu carbon nhất tìm thấy trong Vũ trụ và có thể hình thành trong môi trường quanh các sao khổng lồ đỏ hoặc các đám mây bụi và khí liên sao.[81]
Xem thêm
[sửa | sửa mã nguồn]- RNA origami
- Cấu trúc phân tử sinh học
- Đại phân tử
- DNA
- Transcriptome
Tham khảo
[sửa | sửa mã nguồn]- ^ "RNA: The Versatile Molecule". University of Utah. 2015. Bản gốc lưu trữ ngày 20 tháng 7 năm 2016. Truy cập ngày 3 tháng 6 năm 2018.
- ^ "Nucleotides and Nucleic Acids" (PDF). University of California, Los Angeles. Bản gốc (PDF) lưu trữ ngày 23 tháng 9 năm 2015. Truy cập ngày 3 tháng 6 năm 2018.
- ^ Shukla RN (ngày 30 tháng 6 năm 2014). Analysis of Chromosomes. ISBN 9789384568177.[liên kết hỏng]
- ^ a b c Berg JM, Tymoczko JL, Stryer L (2002). Biochemistry (ấn bản thứ 5). WH Freeman and Company. tr. 118–19, 781–808. ISBN 0-7167-4684-0. OCLC 179705944.
- ^ Tinoco I, Bustamante C (tháng 10 năm 1999). "How RNA folds". Journal of Molecular Biology. Quyển 293 số 2. tr. 271–81. doi:10.1006/jmbi.1999.3001. PMID 10550208.
- ^ Higgs PG (tháng 8 năm 2000). "RNA secondary structure: physical and computational aspects". Quarterly Reviews of Biophysics. Quyển 33 số 3. tr. 199–253. doi:10.1017/S0033583500003620. PMID 11191843.
- ^ a b Nissen P, và đồng nghiệp (tháng 8 năm 2000). "The structural basis of ribosome activity in peptide bond synthesis". Science. Quyển 289 số 5481. tr. 920–30. Bibcode:2000Sci...289..920N. doi:10.1126/science.289.5481.920. PMID 10937990.
- ^ a b Lee JC, Gutell RR (tháng 12 năm 2004). "Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs". Journal of Molecular Biology. Quyển 344 số 5. tr. 1225–49. doi:10.1016/j.jmb.2004.09.072. PMID 15561141.
- ^ Barciszewski J, Frederic B, Clark C (1999). RNA biochemistry and biotechnology. Springer. tr. 73–87. ISBN 0-7923-5862-7. OCLC 52403776.
- ^ Salazar M, và đồng nghiệp (tháng 4 năm 1993). "The DNA strand in DNA.RNA hybrid duplexes is neither B-form nor A-form in solution". Biochemistry. Quyển 32 số 16. tr. 4207–15. doi:10.1021/bi00067a007. PMID 7682844.
- ^ Sedova A, Banavali NK (tháng 10 năm 2015). "RNA approaches the B-form in stacked single strand dinucleotide contexts". Biopolymers. Quyển 105 số 2. tr. 65–82. doi:10.1002/bip.22750. PMID 26443416.
- ^ Hermann T, Patel DJ (tháng 3 năm 2000). "RNA bulges as architectural and recognition motifs". Structure. Quyển 8 số 3. tr. R47–54. doi:10.1016/S0969-2126(00)00110-6. PMID 10745015.
- ^ Mikkola S, và đồng nghiệp (1999). "The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group". Perkin transactions 2. Số 8. tr. 1619–26. doi:10.1039/a903691a.
- ^ Jankowski JA, Polak JM (1996). Clinical gene analysis and manipulation: Tools, techniques and troubleshooting. Cambridge University Press. tr. 14. ISBN 0-521-47896-0. OCLC 33838261.
- ^ Yu Q, Morrow CD (tháng 5 năm 2001). "Identification of critical elements in the tRNA acceptor stem and T(Psi)C loop necessary for human immunodeficiency virus type 1 infectivity". Journal of Virology. Quyển 75 số 10. tr. 4902–6. doi:10.1128/JVI.75.10.4902-4906.2001. PMC 114245. PMID 11312362.
- ^ Elliott MS, Trewyn RW (tháng 2 năm 1984). "Inosine biosynthesis in transfer RNA by an enzymatic insertion of hypoxanthine". The Journal of Biological Chemistry. Quyển 259 số 4. tr. 2407–10. PMID 6365911.
- ^ Cantara WA, và đồng nghiệp (tháng 1 năm 2011). "The RNA Modification Database, RNAMDB: 2011 update". Nucleic Acids Research. Quyển 39 số Database issue. tr. D195–201. doi:10.1093/nar/gkq1028. PMC 3013656. PMID 21071406.
- ^ Söll D, RajBhandary U (1995). TRNA: Structure, biosynthesis, and function. ASM Press. tr. 165. ISBN 1-55581-073-X. OCLC 183036381.
- ^ Kiss T (tháng 7 năm 2001). "Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs". The EMBO Journal. Quyển 20 số 14. tr. 3617–22. doi:10.1093/emboj/20.14.3617. PMC 125535. PMID 11447102.
- ^ King TH, và đồng nghiệp (tháng 2 năm 2003). "Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center". Molecular Cell. Quyển 11 số 2. tr. 425–35. doi:10.1016/S1097-2765(03)00040-6. PMID 12620230.
- ^ Mathews DH, và đồng nghiệp (tháng 5 năm 2004). "Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure". Proceedings of the National Academy of Sciences of the United States of America. Quyển 101 số 19. tr. 7287–92. Bibcode:2004PNAS..101.7287M. doi:10.1073/pnas.0401799101. PMC 409911. PMID 15123812.
- ^ Tan ZJ, Chen SJ (tháng 7 năm 2008). "Salt dependence of nucleic acid hairpin stability". Biophysical Journal. Quyển 95 số 2. tr. 738–52. Bibcode:2008BpJ....95..738T. doi:10.1529/biophysj.108.131524. PMC 2440479. PMID 18424500.
- ^ Vater A, Klussmann S (tháng 1 năm 2015). "Turning mirror-image oligonucleotides into drugs: the evolution of Spiegelmer(®) therapeutics". Drug Discovery Today. Quyển 20 số 1. tr. 147–55. doi:10.1016/j.drudis.2014.09.004. PMID 25236655.
- ^ Nudler E, Gottesman ME (tháng 8 năm 2002). "Transcription termination and anti-termination in E. coli". Genes to Cells. Quyển 7 số 8. tr. 755–68. doi:10.1046/j.1365-2443.2002.00563.x. PMID 12167155.
- ^ Hansen JL, và đồng nghiệp (tháng 8 năm 1997). "Structure of the RNA-dependent RNA polymerase of poliovirus". Structure. Quyển 5 số 8. tr. 1109–22. doi:10.1016/S0969-2126(97)00261-X. PMID 9309225.
- ^ Ahlquist P (tháng 5 năm 2002). "RNA-dependent RNA polymerases, viruses, and RNA silencing". Science. Quyển 296 số 5571. tr. 1270–3. Bibcode:2002Sci...296.1270A. doi:10.1126/science.1069132. PMID 12016304.
- ^ a b c Cooper GC, Hausman RE (2004). The Cell: A Molecular Approach (ấn bản thứ 3). Sinauer. tr. 261–76, 297, 339–44. ISBN 0-87893-214-3. OCLC 174924833.
- ^ Mattick JS, Gagen MJ (tháng 9 năm 2001). "The evolution of controlled multitasked gene networks: the role of introns and other noncoding RNAs in the development of complex organisms". Molecular Biology and Evolution. Quyển 18 số 9. tr. 1611–30. doi:10.1093/oxfordjournals.molbev.a003951. PMID 11504843.
- ^ Mattick JS (tháng 11 năm 2001). "Non-coding RNAs: the architects of eukaryotic complexity". EMBO Reports. Quyển 2 số 11. tr. 986–91. doi:10.1093/embo-reports/kve230. PMC 1084129. PMID 11713189.
- ^ Mattick JS (tháng 10 năm 2003). "Challenging the dogma: the hidden layer of non-protein-coding RNAs in complex organisms" (PDF). BioEssays. Quyển 25 số 10. tr. 930–9. doi:10.1002/bies.10332. PMID 14505360. Bản gốc lưu trữ ngày 6 tháng 3 năm 2009.
- ^ Mattick JS (tháng 10 năm 2004). "The hidden genetic program of complex organisms". Scientific American. Quyển 291 số 4. tr. 60–7. doi:10.1038/scientificamerican1004-60. PMID 15487671. Bản gốc lưu trữ ngày 20 tháng 9 năm 2017. Truy cập ngày 3 tháng 6 năm 2018.
- ^ a b c Wirta W (2006). Mining the transcriptome – methods and applications. Stockholm: School of Biotechnology, Royal Institute of Technology. ISBN 91-7178-436-5. OCLC 185406288.
- ^ Rossi JJ (tháng 7 năm 2004). "Ribozyme diagnostics comes of age". Chemistry & Biology. Quyển 11 số 7. tr. 894–5. doi:10.1016/j.chembiol.2004.07.002. PMID 15271347.
- ^ Storz G (tháng 5 năm 2002). "An expanding universe of noncoding RNAs". Science. Quyển 296 số 5571. tr. 1260–3. Bibcode:2002Sci...296.1260S. doi:10.1126/science.1072249. PMID 12016301.
- ^ Fatica A, Bozzoni I (tháng 1 năm 2014). "Long non-coding RNAs: new players in cell differentiation and development". Nature Reviews. Genetics. Quyển 15 số 1. tr. 7–21. doi:10.1038/nrg3606. PMID 24296535.
- ^ Chen Q, Yan, và đồng nghiệp (tháng 1 năm 2016). "Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder". Science. Quyển 351 số 6271. tr. 397–400. Bibcode:2016Sci...351..397C. doi:10.1126/science.aad7977. PMID 26721680.
- ^ Wei H, Zhou, và đồng nghiệp (2013). "Profiling and identification of small rDNA-derived RNAs and their potential biological functions". PLOS One. Quyển 8 số 2. tr. e56842. Bibcode:2013PLoSO...856842W. doi:10.1371/journal.pone.0056842. PMC 3572043. PMID 23418607.{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết)
- ^ Gueneau de Novoa P, Williams KP (tháng 1 năm 2004). "The tmRNA website: reductive evolution of tmRNA in plastids and other endosymbionts". Nucleic Acids Research. Quyển 32 số Database issue. tr. D104–8. doi:10.1093/nar/gkh102. PMC 308836. PMID 14681369.
- ^ Carthew RW, Sontheimer EJ (tháng 2 năm 2009). "Origins and Mechanisms of miRNAs and siRNAs". Cell. Quyển 136 số 4. tr. 642–55. doi:10.1016/j.cell.2009.01.035. PMC 2675692. PMID 19239886.
- ^ Liang KH, Yeh CT (tháng 5 năm 2013). "A gene expression restriction network mediated by sense and antisense Alu sequences located on protein-coding messenger RNAs". BMC Genomics. Quyển 14. tr. 325. doi:10.1186/1471-2164-14-325. PMC 3655826. PMID 23663499.{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết)
- ^ Wu L, Belasco JG (tháng 1 năm 2008). "Let me count the ways: mechanisms of gene regulation by miRNAs and siRNAs". Molecular Cell. Quyển 29 số 1. tr. 1–7. doi:10.1016/j.molcel.2007.12.010. PMID 18206964.
- ^ Matzke MA, Matzke AJ (tháng 5 năm 2004). "Planting the seeds of a new paradigm". PLoS Biology. Quyển 2 số 5. tr. E133. doi:10.1371/journal.pbio.0020133. PMC 406394. PMID 15138502.{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết)
- ^ Vazquez F, và đồng nghiệp (tháng 10 năm 2004). "Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs". Molecular Cell. Quyển 16 số 1. tr. 69–79. doi:10.1016/j.molcel.2004.09.028. PMID 15469823.
- ^ Watanabe, Toshiaki; Totoki, Yasushi; Toyoda, Atsushi; Kaneda, Masahiro; Kuramochi-Miyagawa, Satomi; Obata, Yayoi; Chiba, Hatsune; Kohara, Yuji; Kono, Tomohiro (ngày 22 tháng 5 năm 2008). "Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes". Nature. Quyển 453 số 7194. tr. 539–543. doi:10.1038/nature06908. ISSN 1476-4687. PMID 18404146.
- ^ Sontheimer EJ, Carthew RW (tháng 7 năm 2005). "Silence from within: endogenous siRNAs and miRNAs". Cell. Quyển 122 số 1. tr. 9–12. doi:10.1016/j.cell.2005.06.030. PMID 16009127.
- ^ Doran G (2007). "RNAi – Is one suffix sufficient?". Journal of RNAi and Gene Silencing. Quyển 3 số 1. tr. 217–19. Bản gốc lưu trữ ngày 16 tháng 7 năm 2007.
- ^ Pushparaj PN, và đồng nghiệp (tháng 1 năm 2008). "RNAi and RNAa--the yin and yang of RNAome". Bioinformation. Quyển 2 số 6. tr. 235–7. doi:10.6026/97320630002235. PMC 2258431. PMID 18317570.
- ^ Horwich MD, và đồng nghiệp (tháng 7 năm 2007). "The Drosophila RNA methyltransferase, DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC". Current Biology. Quyển 17 số 14. tr. 1265–72. doi:10.1016/j.cub.2007.06.030. PMID 17604629.
- ^ Girard A, và đồng nghiệp (tháng 7 năm 2006). "A germline-specific class of small RNAs binds mammalian Piwi proteins". Nature. Quyển 442 số 7099. tr. 199–202. Bibcode:2006Natur.442..199G. doi:10.1038/nature04917. PMID 16751776.
- ^ Horvath P, Barrangou R (tháng 1 năm 2010). "CRISPR/Cas, the immune system of bacteria and archaea". Science. Quyển 327 số 5962. tr. 167–70. Bibcode:2010Sci...327..167H. doi:10.1126/science.1179555. PMID 20056882.
- ^ Wagner EG, Altuvia S, Romby P (2002). "Antisense RNAs in bacteria and their genetic elements". Advances in Genetics. Advances in Genetics. Quyển 46. tr. 361–98. doi:10.1016/S0065-2660(02)46013-0. ISBN 9780120176465. PMID 11931231.
- ^ Developmental Biology (ấn bản thứ 7). Sinauer. 2003. tr. 101–3. ISBN 0-87893-258-5. OCLC 154656422. {{Chú thích sách}}: Đã bỏ qua tham số không rõ |authors= (trợ giúp)
- ^ Amaral PP, Mattick JS (tháng 8 năm 2008). "Noncoding RNA in development". Mammalian Genome. Quyển 19 số 7–8. tr. 454–92. doi:10.1007/s00335-008-9136-7. PMID 18839252.
- ^ Heard E, và đồng nghiệp (tháng 6 năm 1999). "Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells". Proceedings of the National Academy of Sciences of the United States of America. Quyển 96 số 12. tr. 6841–6. Bibcode:1999PNAS...96.6841H. doi:10.1073/pnas.96.12.6841. PMC 22003. PMID 10359800.
- ^ Batey RT (tháng 6 năm 2006). "Structures of regulatory elements in mRNAs". Current Opinion in Structural Biology. Quyển 16 số 3. tr. 299–306. doi:10.1016/j.sbi.2006.05.001. PMID 16707260.
- ^ Scotto L, Assoian RK (tháng 6 năm 1993). "A GC-rich domain with bifunctional effects on mRNA and protein levels: implications for control of transforming growth factor beta 1 expression". Molecular and Cellular Biology. Quyển 13 số 6. tr. 3588–97. PMC 359828. PMID 8497272. Bản gốc lưu trữ ngày 24 tháng 9 năm 2019. Truy cập ngày 3 tháng 6 năm 2018.
- ^ Steitz TA, Steitz JA (tháng 7 năm 1993). "A general two-metal-ion mechanism for catalytic RNA". Proceedings of the National Academy of Sciences of the United States of America. Quyển 90 số 14. tr. 6498–502. Bibcode:1993PNAS...90.6498S. doi:10.1073/pnas.90.14.6498. PMC 46959. PMID 8341661.
- ^ Xie J, và đồng nghiệp (tháng 1 năm 2007). "Sno/scaRNAbase: a curated database for small nucleolar RNAs and cajal body-specific RNAs". Nucleic Acids Research. Quyển 35 số Database issue. tr. D183–7. doi:10.1093/nar/gkl873. PMC 1669756. PMID 17099227.
- ^ Omer AD, và đồng nghiệp (tháng 5 năm 2003). "RNA-modifying machines in archaea". Molecular Microbiology. Quyển 48 số 3. tr. 617–29. doi:10.1046/j.1365-2958.2003.03483.x. PMID 12694609.
- ^ Cavaillé J, và đồng nghiệp (tháng 10 năm 1996). "Targeted ribose methylation of RNA in vivo directed by tailored antisense RNA guides". Nature. Quyển 383 số 6602. tr. 732–5. Bibcode:1996Natur.383..732C. doi:10.1038/383732a0. PMID 8878486.
- ^ Kiss-László Z, và đồng nghiệp (tháng 6 năm 1996). "Site-specific ribose methylation of preribosomal RNA: a novel function for small nucleolar RNAs". Cell. Quyển 85 số 7. tr. 1077–88. doi:10.1016/S0092-8674(00)81308-2. PMID 8674114.
- ^ Daròs JA, Elena SF, Flores R (tháng 6 năm 2006). "Viroids: an Ariadne's thread into the RNA labyrinth". EMBO Reports. Quyển 7 số 6. tr. 593–8. doi:10.1038/sj.embor.7400706. PMC 1479586. PMID 16741503.
- ^ Kalendar R, và đồng nghiệp (tháng 3 năm 2004). "Large retrotransposon derivatives: abundant, conserved but nonautonomous retroelements of barley and related genomes". Genetics. Quyển 166 số 3. tr. 1437–50. doi:10.1534/genetics.166.3.1437. PMC 1470764. PMID 15082561.
- ^ Podlevsky JD, và đồng nghiệp (tháng 1 năm 2008). "The telomerase database". Nucleic Acids Research. Quyển 36 số Database issue. tr. D339–43. doi:10.1093/nar/gkm700. PMC 2238860. PMID 18073191.
- ^ Blevins T, và đồng nghiệp (2006). "Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing". Nucleic Acids Research. Quyển 34 số 21. tr. 6233–46. doi:10.1093/nar/gkl886. PMC 1669714. PMID 17090584.
- ^ Jana S, và đồng nghiệp (tháng 11 năm 2004). "RNA interference: potential therapeutic targets". Applied Microbiology and Biotechnology. Quyển 65 số 6. tr. 649–57. doi:10.1007/s00253-004-1732-1. PMID 15372214.
- ^ Schultz U, Kaspers B, Staeheli P (tháng 5 năm 2004). "The interferon system of non-mammalian vertebrates". Developmental and Comparative Immunology. Quyển 28 số 5. tr. 499–508. doi:10.1016/j.dci.2003.09.009. PMID 15062646.
- ^ Whitehead KA, và đồng nghiệp (2011). "Silencing or stimulation? siRNA delivery and the immune system". Annual Review of Chemical and Biomolecular Engineering. Quyển 2. tr. 77–96. doi:10.1146/annurev-chembioeng-061010-114133. PMID 22432611.
- ^ Dahm R (tháng 2 năm 2005). "Friedrich Miescher and the discovery of DNA". Developmental Biology. Quyển 278 số 2. tr. 274–88. doi:10.1016/j.ydbio.2004.11.028. PMID 15680349.
- ^ Caspersson T, Schultz J (1939). "Pentose nucleotides in the cytoplasm of growing tissues". Nature. Quyển 143 số 3623. tr. 602–3. Bibcode:1939Natur.143..602C. doi:10.1038/143602c0.
- ^ Ochoa S (1959). "Enzymatic synthesis of ribonucleic acid" (PDF). Nobel Lecture.
- ^ Rich A, Davies D (1956). "A New Two-Stranded Helical Structure: Polyadenylic Acid and Polyuridylic Acid". Journal of the American Chemical Society. Quyển 78 số 14. tr. 3548–3549. doi:10.1021/ja01595a086.
- ^ Holley RW, và đồng nghiệp (tháng 3 năm 1965). "Structure of a ribonucleic acid". Science. Quyển 147 số 3664. tr. 1462–5. Bibcode:1965Sci...147.1462H. doi:10.1126/science.147.3664.1462. PMID 14263761.
- ^ Fiers, W.; Contreras, R.; Duerinck, F.; Haegeman, G.; Iserentant, D.; Merregaert, J.; Min Jou, W.; Molemans, F.; Raeymaekers, A. (ngày 8 tháng 4 năm 1976). "Complete nucleotide sequence of bacteriophage MS2 RNA: primary and secondary structure of the replicase gene". Nature. Quyển 260 số 5551. tr. 500–507. doi:10.1038/260500a0. ISSN 0028-0836. PMID 1264203.
- ^ Napoli C, Lemieux C, Jorgensen R (tháng 4 năm 1990). "Introduction of a Chimeric Chalcone Synthase Gene into Petunia Results in Reversible Co-Suppression of Homologous Genes in trans". The Plant Cell. Quyển 2 số 4. tr. 279–289. doi:10.1105/tpc.2.4.279. PMC 159885. PMID 12354959.
- ^ Dafny-Yelin M, và đồng nghiệp (tháng 12 năm 2007). "pSAT RNA interference vectors: a modular series for multiple gene down-regulation in plants". Plant Physiology. Quyển 145 số 4. tr. 1272–81. doi:10.1104/pp.107.106062. PMC 2151715. PMID 17766396.
- ^ Ruvkun G (tháng 10 năm 2001). "Molecular biology. Glimpses of a tiny RNA world". Science. Quyển 294 số 5543. tr. 797–9. doi:10.1126/science.1066315. PMID 11679654.
- ^ Fichou Y, Férec C (tháng 12 năm 2006). "The potential of oligonucleotides for therapeutic applications". Trends in Biotechnology. Quyển 24 số 12. tr. 563–70. doi:10.1016/j.tibtech.2006.10.003. PMID 17045686.
- ^ Siebert S (2006). "Common sequence structure properties and stable regions in RNA secondary structures" (PDF). Dissertation, Albert-Ludwigs-Universität, Freiburg im Breisgau. tr. 1. Bản gốc lưu trữ ngày 9 tháng 3 năm 2012.
- ^ Szathmáry E (tháng 6 năm 1999). "The origin of the genetic code: amino acids as cofactors in an RNA world". Trends in Genetics. Quyển 15 số 6. tr. 223–9. doi:10.1016/S0168-9525(99)01730-8. PMID 10354582.
- ^ Marlaire, Ruth (ngày 3 tháng 3 năm 2015). "NASA Ames Reproduces the Building Blocks of Life in Laboratory". NASA. Bản gốc lưu trữ ngày 5 tháng 3 năm 2015. Truy cập ngày 5 tháng 3 năm 2015.
Liên kết ngoài
[sửa | sửa mã nguồn]- RNA tại Từ điển bách khoa Việt Nam
- RNA (biochemistry) tại Encyclopædia Britannica (bằng tiếng Anh)
- RNA World website Link collection (structures, sequences, tools, journals)
- Nucleic Acid Database Images of DNA, RNA and complexes.
- Anna Marie Pyle's Seminar: RNA Structure, Function, and Recognition Lưu trữ ngày 10 tháng 11 năm 2017 tại Wayback Machine
| |||||||
|---|---|---|---|---|---|---|---|
| Thành phần hợp thành |
|  | |||||
| Acid ribonucleic(biên soạn mã, không biên soạn mã) |
| ||||||
| Acid deoxyribonucleic |
| ||||||
| Chất tương tự |
| ||||||
| Vectơ tách dòng |
| ||||||
| |
|---|---|
| |
| Thành phần then chốt |
|
| Chủ đề chính |
|
| Khảo cổ học di truyền |
|
| Chủ đề liên quan |
|
| |
| |||||||
|---|---|---|---|---|---|---|---|
| Giới thiệu về di truyền học |
|  | |||||
| Phiên mã |
| ||||||
| Dịch mã |
| ||||||
| Biểu hiện gen |
| ||||||
| Người có tầm ảnh hưởng |
| ||||||
| |
|---|---|
| Hệ thống nội màng |
|
| Khung xương tế bào |
|
| Nội cộng sinh |
|
| Cấu trúc nội bào khác |
|
| Cấu trúc ngoại bào |
|
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antifolate(ức chếquá trình chuyển hóa purine,do đó ức chế tổng hợp ADN và ARN) |
| ||||||||||||||||
| Chất ức chế Topoisomerase/quinolone/(ức chế quá trình nhân đôi DNA) |
| ||||||||||||||||
| Chất ức chế ADN yếm khí |
| ||||||||||||||||
| Tổng hợp RNA |
| ||||||||||||||||
| #WHO-EM. ‡Thu hồi trên thị trường. Thử nghiệm lâm sàng: †Pha III. §Chưa bao giờ đến pha III | |||||||||||||||||
| ||||||||
|---|---|---|---|---|---|---|---|---|
| ||||||||
| Tổng quan |
|   | ||||||
| Kỹ thuật |
| |||||||
| ||||||||
| Cơ sở dữ liệu tiêu đề chuẩn | |
|---|---|
| Quốc tế |
|
| Quốc gia |
|
| Khác |
|
Từ khóa » Chiều Của Arn Thông Tin
-
RNA Thông Tin – Wikipedia Tiếng Việt
-
Cấu Tạo Và Chức Năng Của ARN
-
Tổng Quan Về Di Truyền Học - Phiên Bản Dành Cho Chuyên Gia
-
CẤU TRÚC VÀ CHỨC NĂNG CỦA ADN VÀ ARN - Flat World
-
Lý Thuyết ARN Và Quá Trình Phiên Mã Sinh 12
-
Phiên Mã Và Dịch Mã, Trắc Nghiệm Hóa Học Lớp 12
-
2. Phiên Mã Và Dịch Mã - Củng Cố Kiến Thức
-
ARN được Tổng Hợp Từ Mạch Nào Của ADN - Luật Hoàng Phi
-
[PDF] Câu 1: Trong Quá Trình Dịch Mã, Trên Một Phân Tử MARN Thường Có ...
-
Bài Giảng Khái Niệm Về Di Truyền Và Biểu Hiện ... - Health Việt Nam
-
Các Kỹ Sư Tạo Ra được Dạng Hít Của ARN Thông Tin Cấp Nano
-
[PDF] CÁC CÔNG THỨC TÍNH TOÁN - VNUF2
-
[PDF] Phiên Mã Và Dịch Mã.
-
[PDF] KỲ THI KIỂM TRA NĂNG LỰC TRƯỜNG ĐẠI HỌC QUỐC TẾ BÀI THI