Stable C0T-1 Repeat RNA Is Abundant And Is Associated ... - PubMed
Abstract
Recent studies recognize a vast diversity of noncoding RNAs with largely unknown functions, but few have examined interspersed repeat sequences, which constitute almost half our genome. RNA hybridization in situ using C0T-1 (highly repeated) DNA probes detects surprisingly abundant euchromatin-associated RNA comprised predominantly of repeat sequences (C0T-1 RNA), including LINE-1. C0T-1-hybridizing RNA strictly localizes to the interphase chromosome territory in cis and remains stably associated with the chromosome territory following prolonged transcriptional inhibition. The C0T-1 RNA territory resists mechanical disruption and fractionates with the nonchromatin scaffold but can be experimentally released. Loss of repeat-rich, stable nuclear RNAs from euchromatin corresponds to aberrant chromatin distribution and condensation. C0T-1 RNA has several properties similar to XIST chromosomal RNA but is excluded from chromatin condensed by XIST. These findings impact two "black boxes" of genome science: the poorly understood diversity of noncoding RNA and the unexplained abundance of repetitive elements.
Copyright © 2014 Elsevier Inc. All rights reserved.
PubMed Disclaimer
Figures
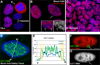
Figure 1. CoT-1 RNA is expressed in…
Figure 1. CoT-1 RNA is expressed in all mammalian cells examined
All blue signals are…
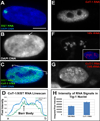
Figure 2. CoT-1 RNA appears highly abundant
Figure 2. CoT-1 RNA appears highly abundant
A) XIST RNA paints interphase X-chromosome territory. Scale…
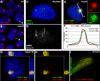
Figure 3. CoT-1 RNA localizes to the…
Figure 3. CoT-1 RNA localizes to the chromosome similar to XIST RNA
A) XIST RNA…
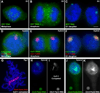
Figure 4. The majority of RNA associated…
Figure 4. The majority of RNA associated with interphase chromosomes is repeat RNA, which is…
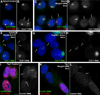
Figure 5. CoT-1 RNA localization is very…
Figure 5. CoT-1 RNA localization is very stable under transcriptional inhibition
A–B) CoT-1 RNA is…
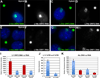
Figure 6. RNA from the 3’ end…
Figure 6. RNA from the 3’ end of L1 is a large component of the…
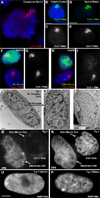
Figure 7. CoT-1 RNA associates with the…
Figure 7. CoT-1 RNA associates with the nuclear scaffold and Cot-1 RNA loss is coincident…
Comment in
- Interphase chromatin LINEd with RNA. Nozawa RS, Gilbert N. Nozawa RS, et al. Cell. 2014 Feb 27;156(5):864-5. doi: 10.1016/j.cell.2014.02.005. Cell. 2014. PMID: 24581484
References
-
- Aalto AP, Pasquinelli AE. Small non-coding RNAs mount a silent revolution in gene expression. Current opinion in cell biology. 2012;24:333–340. - PMC - PubMed
-
- Bensaude O. Inhibiting eukaryotic transcription: Which compound to choose? How to evaluate its activity? Transcription. 2011;2:103–108. - PMC - PubMed
-
- Britten RJ, Kohne DE. Repeated sequences in DNA: Hundreds of thousands of copies of DNA sequences have been incorporated into the genomes of higher organisms. Science. 1968;161:529–540. - PubMed
-
- Bynum JW, Volkin E. Chromatin-associated RNA: differential extraction and characterization. Biochimica et biophysica acta. 1980;607:304–318. - PubMed
-
- Chaumeil J, Le Baccon P, Wutz A, Heard E. A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes Dev. 2006;20:2223–2237. - PMC - PubMed
Publication types
- Research Support, N.I.H., Extramural Actions
- Search in PubMed
- Search in MeSH
- Add to Search
MeSH terms
- Animals Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Cell Nucleus / chemistry Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Chromosomes, Mammalian / chemistry* Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Euchromatin / chemistry* Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Humans Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Hybrid Cells Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Interphase* Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Long Interspersed Nucleotide Elements Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Mice Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- RNA, Untranslated / analysis* Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- RNA, Untranslated / genetics Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Repetitive Sequences, Nucleic Acid Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- Transcription, Genetic Actions
- Search in PubMed
- Search in MeSH
- Add to Search
Substances
- Euchromatin Actions
- Search in PubMed
- Search in MeSH
- Add to Search
- RNA, Untranslated Actions
- Search in PubMed
- Search in MeSH
- Add to Search
Grants and funding
- F32 CA154086/CA/NCI NIH HHS/United States
- 1F32CA154086/CA/NCI NIH HHS/United States
- GM107604/GM/NIGMS NIH HHS/United States
- R01 GM107604/GM/NIGMS NIH HHS/United States
- GM053234/GM/NIGMS NIH HHS/United States
- R01 GM053234/GM/NIGMS NIH HHS/United States
- S10RR027897/RR/NCRR NIH HHS/United States
- S10 RR027897/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
- Elsevier Science
- Europe PubMed Central
- PubMed Central
Other Literature Sources
- H1 Connect - Access expert opinions and insights on biomedical research.
- The Lens - Patent Citations Database
- scite Smart Citations
Molecular Biology Databases
- NIAID Data Ecosystem - Find datasets on Infectious and Immune-mediated Diseases
Từ khóa » C0t
-
C0t Curve - Oxford Reference
-
A Rapid Procedure For The Isolation Of C0t-1 DNA From Plants - PubMed
-
C0T Ligand Summary Page - RCSB PDB
-
C0T | THE FINEST TASTE IN MODERN CUISINE - LUBUDS
-
Stable C0T-1 Repeat RNA Is Abundant And Is Associated With ...
-
LM3S5R31-IQC80-C0T Texas Instruments | Mouser Singapore
-
[PDF] A Rapid Procedure For The Isolation Of Cot-1 DNA From Plants
-
Chemical Components In The PDB - EMBL-EBI