Synap Hóa Học – Wikipedia Tiếng Việt
Có thể bạn quan tâm

Synap hóa học là nơi tiếp xúc giữa hai nơron, mà thông qua đó cho phép các nơron truyền đi thông tin qua lại lẫn nhau (chỉ một chiều). Synap không chỉ hiện diện ở các tế bào thần kinh, mà còn ở tế bào cơ và tế bào tuyến bằng việc các nơron tạo synap với các tế bào đó. Nhờ việc tạo synap này mới dẫn đến hình thành các mạch nơron trong hệ thống thần kinh trung ương. Với quy mô của sự phức tạp ở synap, và minh chứng cho sự tính toán hoàn hảo của hệ thống sinh học này, tất cả đều là nền tảng cho sự hình thành tư duy và cả nhận thức. Chúng cung cấp cho hệ thần kinh khả năng kết nối và kiểm soát tất cả các hệ thống khác của cơ thể.
Tại synap hóa học, một nơron giải phóng các phân tử dẫn truyền tín hiệu vào nơron kế cận với nó qua khe synap (khoảng trống nhỏ). Bao synap là những bao nhỏ chứa chất dẫn truyền thần kinh trong đó, được giải phóng vào khe synap bởi cơ chế xuất bào. Những phân tử truyền tin này bắt đầu gắn vào các thụ thể trên màng tế bào sau synap. Và cuối cùng, khi đã hoàn thành xong nhiệm vụ truyền tin của mình, các chất dẫn truyền thần kinh này bị tiêu biến ngay tại synap bởi các cơ chế của hệ thần kinh bao gồm có thoái hóa bởi enzyme hoặc là tái hấp thu qua các kênh protein vận chuyển xuyên màng, ở màng tế bào trước synap hay là ở một số loại tế bào thần kinh đệm (neuroglia) nhằm chấm dứt hoạt động của các phân tử truyền tin này.
Bộ não của người trưởng thành ước tính có khoảng 1014 đến 5 1014 (100–500 nghìn tỷ) synap.[1] Mỗi milimét khối lớp vỏ não chứa xấp xỉ khoảng 1 tỷ (theo quy ước số ngắn, tức là 109) synap.[2] Phân tích toàn bộ lớp vỏ não đã được tách ra riêng biệt cho thấy có rất nhiều synap và ước tính giá trị khoảng 0.15 nghìn triệu triệu (150 nghìn tỷ).[3]

Từ ngữ "synap" được đề xướng bởi ngài Charles Scott Sherrington vào năm 1897.[4] Synap hóa không chỉ là dạng duy nhất tồn tại ở sinh vật sống: ngoài ra còn có synap điện và synap miễn dịch. Tuy nhiên là nếu không dựa theo tiêu chuẩn đặc biệt, thì thuật ngữ "synap" thường được xem là synap hóa học.
Cấu trúc Synap
[sửa | sửa mã nguồn]| Cấu trúc điển hình của một synap hóa học |
|---|
 Vùng đặc sau synap Cổng kênh điện thế Ca++ Bọc synap Chất dẫn truyền thần kinh Kênh protein tái hấp thu Thụ thể Cúc tận cùng Khe synap Sợi nhánh Vùng đặc sau synap Cổng kênh điện thế Ca++ Bọc synap Chất dẫn truyền thần kinh Kênh protein tái hấp thu Thụ thể Cúc tận cùng Khe synap Sợi nhánh |
Synap thực hiện chức năng của nó chính là kết nối giữa các nơron lại với nhau, hoặc giữa nơron với các loại tế bào khác.[5][6] Một nơron điển hình sẽ tạo vài nghìn synap, một số nơron khác có thể nhiều hơn hoặc ít hơn số đó.[7] Hầu hết synap chúng kết nối sợi trục (axons) với sợi nhánh (dendrites),[8][9] nhưng đồng thời cũng tồn tại các dạng kết nối khác, gồm có sợi trục với thân tế bào,[10][11] sợi trục với sợi trục,[10][11] và sợi nhánh với sợi nhánh.[9] Nhìn chung các synap này quá nhỏ để có thể nhận diện rõ dưới kính hiển vi quang học và chỉ có thể thấy rõ điểm mà hai màng tế bào tiếp xúc nhau, nhưng khi ta nhìn bằng chiếc kính hiển vi điện tử thì có thể thấy rõ được mọi thành phần tế bào của nó.
| Khác biệt giữa nơron trước synap và nơron sau synap[12] |
|---|
 Khe synap { ↓ Từ nơron trước synap Truyền dẫn các chất dẫn truyền thần kinh qua synap ↓ Đến nơron sau synap Kênh vận chuyển chất dẫn truyền Thụ thể chất dẫn truyền Khe synap { ↓ Từ nơron trước synap Truyền dẫn các chất dẫn truyền thần kinh qua synap ↓ Đến nơron sau synap Kênh vận chuyển chất dẫn truyền Thụ thể chất dẫn truyền |
| "Nơron này kết nối với nơron kia thông qua synap. Dòng tín hiệu di chuyển chỉ theo một chiều, đi từ nơron trước synap đến nơron sau synap qua khe synap, đồng thời khe synap cũng đóng vai trò là trạm dừng của các đường tín hiệu".[12] Hướng của dòng tín hiệu giúp xác định xem là nơron nào là trước synap và nơron nào là sau synap.[12] |
Các synap hóa học tạo điều kiện cho việc truyền dẫn thông tin tế bào trực tiếp từ tế bào trước synap đến tế bào sau synap, và vì thế nên cả cấu trúc và chức năng của nó mang tính bất đối xứng. Ở tận cùng sợi trục trước synap, hay còn gọi là cúc tận cùng, là khu vực chuyên biệt chứa các chất dẫn truyền thần kinh (neurotransmitter), các phân tử dẫn truyền này được gói trong cấu trúc hình cầu nhỏ có thể gắn với màng tế bào thực hiện quá trình xuất bào được gọi là bọc synap (cũng như là nhiều cấu trúc và bào quan khác đóng vai trò hỗ trợ, như là ty thể và mạng lưới nội chất). Các bọc synap (synaptic vesicles) được giải phóng ngay tại màng tế bào trước synap tại vùng đặc biệt được gọi là vùng hoạt động (active zones).
Ở vị trí đối diện các cúc tận cùng (axon terminals) chính là vùng tế bào sau synap chứa các thụ thể trên đó; và vùng sau synap có thể hiện diện ở sợi nhánh hoặc là thân tế bào (soma). Ngay tại màng sau synap, kế tiếp có các chuỗi phức hợp protein chúng liên kết với nhau tinh vi và rất phức tạp chứa trong vùng đặc biệt với tên gọi vùng đặc sau synap (PSD).
Các protein trong vùng đặc sau synap thực hiện nhiệm vụ bắm chặt lấy các thụ thể chất dẫn truyền thần kinh và vận chuyển nó đến màng tế bào sau synap, cũng như là vai trò điều phối hoạt động của các thụ thể này. Các thụ thể và vùng đặc sau synap thường thấy nằm ở gai sợi nhánh (dendritic spine), một cấu trúc đặc biệt nhô ra từ thân sợi nhánh.
Các synap hoặc là mang tính đối xứng hoặc là bất đối xứng. Khi thực hiện phân tích các cấu trúc synap dưới kính hiển vi điện tử, synap bất đối xứng được đặc trưng bởi có các bọc tròn synap ở tế bào trước synap, và vùng đặc sau synap chứa diện tích lớn. Các synap bất đối xứng điển hình là các synap kích thích. Và ngược lại các synap đối xứng có các bọc synap dẹt phẳng và dài ra, vùng đặc sau synap không có hoặc chiếm diện tích rất nhỏ. Các synap đối xứng thường là các synap ức chế.
Khe synap — cũng có thể gọi là khoảng trống synap — là khoảng trống giữa các tế bào trước synap và tế bào sau synap và nó rộng khoảng 20 nm (0,02 μ).[12] Với thể tích của khe synap nhỏ như thế, cho phép chất dẫn truyền thần kinh di chuyển nhanh qua khe đến thụ thể tạo kích thích nhanh, và cũng như là ức chế nhanh chóng khi ngừng truyền đạt chất dẫn truyền.[13]
Tự synap (hóa hoặc điện) là hiện tượng cúc tận cùng của một nơron tự tạo synap với chính sợi nhánh của nơron đó.
Quá trình truyền tin diễn ra ở synap hóa học
[sửa | sửa mã nguồn]Tổng quan
[sửa | sửa mã nguồn]Dưới đây là một chuỗi các sự kiện đã được tóm lược lại một cách ngắn gọn và đơn giản hóa diễn ra trong quá trình truyền tin đi từ nơron trước synap đến nơron sau synap. Mỗi bước sẽ được giải thích chi tiết và cụ thể hơn ở phía dưới. Đặc biệt chú ý là chỉ ngoại trừ bước cuối, toàn bộ quá trình còn lại vận hành chỉ trong khoảng thời gian vài trăm μs, đó là khi ta xét đến synap với khả năng dẫn truyền nhanh nhất.[14]
- Quá trình khởi đầu bằng việc sóng kích thích điện hóa được gọi dưới cái tên điện thế hoạt động di chuyển dọc theo màng tế bào trước synap để cuối cùng đến synap.
- Khi đến synap, điện thế hoạt động làm khử cực màng tế bào tại synap dẫn đến mở các kênh protein Ca2+ tăng tính thấm cho ion calci.
- Các ion calci lưu chuyển vào bên trong màng tế bào trước synap, kết quả làm tăng nồng độ calci nội bào.
- Nồng độ calci tăng cao làm hoạt hóa một loạt protein nhạy calci gắn vào các bọc chứa chất dẫn truyền thần kinh.
- Những protein này chúng thay đổi hình dạng, tác động đến màng của các bọc synap "neo đậu" gần đó hợp nhất với lại màng tế bào trước synap, dẫn đến việc giải phóng các chất dẫn truyền thần kinh vào khe synap, khoảng không gian hẹp giữa màng tế bào trước synap và sau synap.
- Chất dẫn truyền thần kinh khuếch tán vào khe synap. Sau đó chúng gắn vào các thụ thể đặc hiệu nằm trên màng tế bào sau synap. Không phải tất cả phân tử truyền đạt khi được giải phóng ra từ bọc synap đều gắn vào thụ thể hết, một số ít phân tử không đến được thụ thể và gắn vào nó.
- Khi chất truyền đạt thần kinh gắn vào thụ thể sẽ hoạt hóa nó theo nhiều cách thức đặc trưng cho tính chất cũng như là cấu trúc hình thể của chính loại thụ thể đó. Một vài dạng hoạt hóa điển hình sẽ được mô tả chi tiết hơn ở bên dưới. Đối với tất cả các cơ chế hoạt hóa thụ thể, điều cốt lõi không ngoài khác hơn đó chính là làm vận hành tế bào sau synap.
- Do chuyển động nhiệt của các phân tử, sự vận động của nguyên tử, làm chuyển dời vị trí cân bằng của khối tinh thể rắn, các phân tử truyền đạt này rồi cuối cùng cũng sẽ bị bật ra khỏi các thụ thể và trôi dạt vào chất nền ngoại bào.
- Chất truyền đạt thần kinh hoặc là bị tái hấp thu vào tế bào trước synap, và sau đó tái đóng gói cho lần giải phóng tiếp theo, hoặc là chúng đi vào quá trình chuyển hóa.
Giải phóng các chất dẫn truyền thần kinh
[sửa | sửa mã nguồn]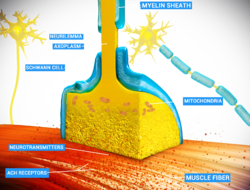
Xung động thần kinh (hay điện thế hoạt động) xuất hiện và di chuyển dọc theo sợi trục gây ra quá trình giải phóng các chất dẫn truyền thần kinh thông qua cơ chế xuất bào. Trong tận cùng sợi trục trước synap, các bọc mang các chất dẫn truyền thần kinh khu trú ở gần màng tế bào. Điện thế hoạt động khi lan truyền đến các cúc tận cùng, hoạt hóa các kênh cổng điện thế calci lưu thông dòng chảy ion calci vào bên trong tế bào.[15] Các ion calci lúc này chúng sẽ tiến hành gắn vào các protein synaptotagmin hiện diện trên màng của các bọc synap, như thế làm cho các bọc này hợp nhất được với màng tế bào trước synap.[16] Quá trình hợp nhất màng này diễn ra theo một cách ngẫu nhiên, do đó dẫn đến việc là quá trình truyền tin qua synap cũng thường xuyên thất bại ở những synap rất nhỏ, và các synap này điển hình có mặt ở hệ thống thần kinh trung ương. Trái ngược lại ở các synap lớn (chẳng hạn như synap thần kinh - cơ), xác suất giải phóng các bọc synap đạt lên tới 100%. Cơ chế vận hành quá trình hợp nhất các bọc synap chính là làm khởi động nên một loạt các protein được biết với tên gọi là SNARE. Nhìn chung là các phức hợp protein này đóng vai trò như chất trung gian điều phối hoạt động hợp nhất màng và cố định vị trí các bọc synap trong vùng hoạt động.[17] Các màng thuộc bọc synap sau khi hợp nhất hóa với màng tế bào sau synap, chúng sẽ được khôi phục trở lại bằng quá trình nhập bào và sau đó được tái sử dụng tham gia quá trình tạo các bọc synap mới.
Tuy cơ chế giải phóng chất dẫn truyền thần kinh bằng việc hòa màng bọc synap chiếm đại đa số diễn ra ở cấp độ thần kinh. Nhưng vẫn có ngoại lệ, đặc biệt đó là thụ thể loại II trên các tế bào nụ vị giác ở động vật có vú. Ở đây chất dẫn truyền thần kinh ATP được giải phóng trực tiếp ngay tại tế bào chất vào qua các cổng kênh điện thế để vào khe synap mà không cần bọc synap cũng như là cơ chế hòa màng như trên.[18]
Gắn vào thụ thể
[sửa | sửa mã nguồn]Các thụ thể ở vị trí đối diện cúc tận cùng trên màng tế bào sau synap để cho các chất dẫn truyền thần kinh gắn vào. Khi các chất truyền đạt gắn vào hoạt hóa thụ thể, nó sẽ đáp ứng bằng một trong hai cách. Thứ nhất, các thụ thể với bản chất là kênh cổng chất gắn sẽ mở trực tiếp ở màng tế bào sau synap, thúc đẩy sự vận động của các ion ra vào tế bào và dẫn đến làm thay đổi cục bộ điện thế màng tế bào. Kết quả của sự thay đổi điện thế đó chính là hình thành điện thế sau synap. Về tổng quan, hiện tượng kích thích (hay hưng phấn) làm tăng dòng chảy ion dương vào nội bào được biết đến chính là quá trình khử cực, và hiện tượng ức chế ngược lại làm tăng dòng chảy ion âm vào nội bào đến mức vượt ngưỡng mức điện thế nghỉ màng tế bào tại thời điểm ban đầu (siêu phân cực). Để mà biết synap nào là ức chế và synap nào là kích thích thì dựa vào tính chất và cấu trúc của kênh ion vận hành dòng chảy ion đặc trưng của kênh ion đó ở màng sau synap, khả năng ứng khớp chức năng của các loại thụ thể đối với từng chất dẫn truyền thần kinh cũng như là có mối tương quan với nó. Thứ hai đó là các thụ thể khi ở dạng hoạt động chúng có thể tác động điện thế màng nhờ vào khả năng kiểm soát quá trình sinh tổng hợp các chất truyền tin thứ hai (second messengers) diễn ra trong nơron sau synap. Những chất truyền tin thứ hai này có thể khuếch đại tín hiệu hưng phấn hay ức chế tùy thuộc vào bản chất của các chất dẫn truyền thần kinh.
Chấm dứt
[sửa | sửa mã nguồn]Sau khi chất dẫn truyền thần kinh đã hoàn thành xong nhiệm vụ của mình là gắn vào thụ thể phân tử, nó cần phải được di dời ngay để tiếp tục tăng cường điện thế sau synap (EPSPs) hoặc là làm giảm điện thế sau synap (IPSPs). Sự biến chuyển các chất truyền đạt này được thực hiện thông qua một hay nhiều quá trình sau:
- Chất dẫn truyền thần kinh có thể bức ra khỏi thụ thể là do tính chuyển động nhiệt của phân tử gây ra bởi cả chính nó và cả thụ thể, tạo điều kiện cho môi trường chất nền ngoại bào bên ngoài nơron nuốt chửng nó (chuyển hóa) hoặc là tái hấp thu.[19]
- Các enzyme bên dưới màng tế bào sau synap chúng có thể làm bất hoạt hay chuyển hóa các chất dẫn truyền thần kinh.
- Các bơm tái hấp thu có thể bơm tích cực các chất dẫn truyền thần kinh trở lại vào tận cùng sợi trục trước synap nhằm tái xử lý sử dụng lại cũng như chờ đợi xung điện thế hoạt động cho lần giải phóng tiếp theo.[19]
Độ mạnh của synap
[sửa | sửa mã nguồn]Ngài Bernard Katz đã ra định nghĩa về độ mạnh của synap được quyết định bởi các yếu tố sau đó là: xác suất giải phóng các sản phẩm sinh hóa trước synap pr (bọc synap), lượng tử số q (khả năng đáp ứng của màng tế bào sau synap với lượng tử trong một bọc synap), và n là số lượng các thụ thể. Thuật ngữ "đơn vị kết nối" trong khoa học thần kinh là ý muốn nói đến các synap chúng kết nối các nơron trước synap và các nơron sau synap lại với nhau, và số lượng của các synap ấy không thể nào tính đếm hết được. Biên độ điện thế sau synap (PSPs) có thể thấp từ 0,4 mV và cao lên đến 20 mV.[20] Và biên độ điện thế sau synap chịu ảnh hưởng bởi các chất dẫn truyền thần kinh khác không chỉ glutamate, và đồng thời có thể thay đổi phụ thuộc vào tần số xung động thần kinh. Những biến đổi độ mạnh synap có thể ngắn hạn (short-term), kéo dài từ nhiều giây cho đến nhiều phút, hoặc là dài hạn (điện thế hóa dài hạn, ĐTHDH), nhiều tiếng và nhiều tháng liền hơn nữa. Quá trình học và nhớ được xem như là gắn liền với sự biến thiên cường độ synap dài hạn, và được thể hiện thông qua cơ chế mềm dẻo của synap (synaptic plasticity).
Ly giải khả năng cảm ứng của thụ thể
[sửa | sửa mã nguồn]Hiện tượng khử nhạy diễn ra ở các thụ thể màng tế bào sau synap chính là khả năng đáp ứng (hay độ nhạy) suy giảm đi đối với cùng mức độ kích thích của cùng một loại chất dẫn truyền thần kinh. Tức điều này có nghĩa là khi nhược hóa một loạt xung điện thế hoạt động tương ứng với tần số cao sẽ dẫn đến hiệu quả truyền tin ở synap giảm đi nhiều lần. Độ mạnh synap sẽ tăng lên khi các synap tiếp nhận kích thích liên tục lặp đi lặp lại nhiều lần (tetanic stimulation), tương ứng các thụ thể sẽ nhạy hóa hơn và các đáp ứng theo sau sẽ trở nên mạnh hơn. Hiện tượng này minh chứng cho việc các synap chúng phụ thuộc rất nhiều vào tần số kích thích. Hệ thống thần kinh đã khéo léo khai thác thuộc tính này cho sự tính toán có mục đích, tức là khử nhạy ở các synap không cần dùng đến hoặc không cần thiết, và nhạy hóa các synap quan trọng bằng quá trình phosphoryl hóa các protein liên quan.
Tính mềm dẻo của synap
[sửa | sửa mã nguồn]Hiệu quả quá trình dẫn truyền thông tin qua synap sẽ tăng nếu như synap đó đã có diễn ra các hoạt động điện thế từ trước, quá trình này được gọi là điện thế hóa, và như thế ngược lại hiệu quả sẽ giảm với quá trình ức chế hóa. Những biến động điện thế này có thể tồn tại dài hạn hoặc ngắn hạn. Dạng mềm dẻo ngắn hạn bao gồm có hiện tượng mỏi synap (synaptic fatigue) và gia tăng điện thế tại synap (synaptic augmentation). Dạng mềm dẻo dài hạn thì gồm có hai cơ chế điển hình nhất đó chính là ức chế hóa dài hạn (long-term depression) và điện thế hóa dài hạn (long-term potentiation). Tính mềm dẻo của synap có thể thể hiện ở synap đơn (homosynaptic plasticity) hoặc là ở nhiều synap khác nhau (heterosynaptic plasticity).
Tính mềm dẻo đơn synap
[sửa | sửa mã nguồn]Tính mềm dẻo đơn synap (còn được gọi là mềm dẻo đồng hướng) thể hiện độ mạnh dẫn truyền của synap thay đổi nhờ vào lịch sử hoạt động điện thế ở tại một synap. Những thay đổi về nồng độ calci nội bào trước synap cũng như là điều chỉnh các thụ thể trên màng, một ví dụ điển hình của dạng truyền tin tự thể (autocrine signaling), cũng hình thành nên tính mềm dẻo đơn synap này. Tính mềm dẻo đơn synap tác động đến số lượng cũng như là tốc độ cung ứng các bọc synap hoặc là ảnh hưởng lên mối quan hệ giữa calci và bọc synap. Về bản chất thì tính mềm dẻo đơn synap có thể xuất hiện ở luôn cả tế bào sau synap. Và kết quả của nó không ngoài việc làm tăng hoặc giảm độ mạnh của synap.
Chẳng hạn như các nơron trong hệ thần kinh giao cảm (SNS), với chất truyền đạt thần kinh là noradrenaline, bên cạnh việc tác động đến thụ thể sau synap, nó còn tác động luôn các thụ thể α2-adrenergic, làm ức chế ngay sự giải phóng noredrenaline.[21] Hiệu quả sinh học này cùng phối hợp với thuốc clonidine ngoại lai để nhằm tăng hiệu quả ức chế trên hệ thống thần kinh giao cảm.
Tính mềm dẻo đa synap
[sửa | sửa mã nguồn]Tính mềm dẻo đa synap (còn được gọi là mềm dẻo dị hướng) là sự thay đổi độ mạnh của synap tùy thuộc vào hoạt động điện thế của các nơron. Và cũng tương tự như trên, tính mềm dẻo làm thay đổi số lượng các bọc synap hoặc tốc độ cung ứng hoặc là mối quan hệ giữa calci và các bọc synap. Thêm vào đó, nó có thể ảnh hưởng trực tiếp đến ion calci bằng việc kiểm soát dòng chảy ion calci vào nội bào. Tính mềm dẻo đa synap cũng có thể hiện diện sau synap, ảnh hưởng lên độ nhạy của thụ thể.
Lấy thêm ví dụ nữa cũng là về nơron trong hệ thống thần kinh giao cảm, không chỉ giải phóng noradrenaline mà còn tạo ra ức chế trên tận cùng nơron trước synap của hệ thần kinh đối giao cảm.[21]
Nhất thể hóa tín hiệu điện thế tại synap
[sửa | sửa mã nguồn]Một cách tổng quát, nếu như kích thích điện thế tại synap đủ mạnh, thì điện thế hoạt động ở nơron trước synap sẽ gây ra hoạt động điện thế tương tự ở tế bào sau synap. Trong nhiều trường hợp điện thế sau synap do kích thích yếu gây ra sẽ chưa thể chạm được đến ngưỡng để hình thành điện thế hoạt động. Khi nhiều điện thế hoạt động ở nhiều nơron trước synap dẫn đến có nhiều kích thích điện cùng một lúc ngay tại synap, hoặc nếu đơn nơron trước synap kích thích với tần số đủ cao, các dòng điện thế sau synap chúng tập hợp lại với nhau và như một phép toán cộng đơn giản. Tổng hợp các điện thế kích thích sau synap có thể chạm đến ngưỡng kích thích làm khởi sinh điện thế hoạt động. Đây chính là hiện tượng cộng kích thích (summation).[22]
Trái lại hoàn toàn thì nơron trước synap giải phóng chất dẫn truyền thần kinh ức chế, điển hình như GABA, có thể gây ra ức chế điện thế ở nơron sau synap (IPSPs), và làm cho điện thế màng tế bào càng xa ngưỡng kích thích hơn nữa, tính kích thích hay hưng phấn giảm đi nhiều và làm cho nơron khó khăn hơn trong việc khởi tạo điện thế hoạt động. Nếu như các hoạt động ức chế điện thế sau synap xen kẽ các hoạt động kích thích sau synap, thì trong nhiều trường hợp sự ức chế đó ngăn chặn nơron khởi sinh điện thế hoạt động. Để giải thích cho điều này, thì có thể nói tín hiệu ra của một nơron phụ thuộc vào nhiều tín hiệu vào của các nơron khác, và mỗi loại nơron có thể sở hữu những mức độ ảnh hưởng khác nhau, cũng như là độ mạnh và loại synap của chính nơron đó. John Carew Eccles đã cùng cộng sự thực hiện một vài công trình nghiên cứu khoa học mang ý nghĩa quan trọng vào đầu những năm 1950 về tính thống nhất hóa kích thích ở synap, nhờ những đóng góp đáng kể cho lĩnh vực khoa học thần kinh này mà ông đã vinh dự được đón nhận giải thưởng Nobel sinh lý học y khoa vào năm 1963.
Truyền dẫn ngoại dịch
[sửa | sửa mã nguồn]Khi chất truyền đạt thần kinh được giải phóng vào synap, đồng nghĩa với việc nồng độ các chất truyền đạt lúc này là cao nhất ngay tại khe synap, và có một lượng chất truyền đạt nhất định khuếch tán ngay lập tức trước khi bị tái hấp thu hoặc bị chuyển hóa tại đó. Nếu các chất khuếch tán ngay không chậm trễ, chúng sẽ hoạt hóa các thụ thể ở gần tại màng tế bào sau synap, hoặc ở xa tại các vị trí synap khác. Phạm vi hoạt động của các chất dẫn truyền thần kinh diễn ra bên ngoài synap được gọi là quá trình truyền dẫn ngoại dịch.[23] Quá trình như thế diễn ra trên phạm vi rộng cũng như là với một mức độ đáng kể, nhưng mà tầm quan trọng về chức năng của nó vẫn còn là vấn đề tranh cãi trong thời gian dài.[24]
Nghiên cứu gần đây cho thấy rằng truyền dẫn ngoại dịch có thể là kiểu tương tác chiếm ưu thế ở một số nơron đặc biệt. Ở lớp vỏ não của động vật có vú, có một loại nơron trung gian với tên gọi là NGF (neurogliaform) nó có thể ức chế các cấu trúc nơron gần đó bằng cách giải phóng chất dẫn truyền thần kinh ức chế đặc trưng GABA vào dịch ngoại bào.[25] Cùng với đó, chất ức chế GABA do các nơron NGF giải phóng ra cũng đồng thời tác dụng lên các tế bào hình sao (astrocytes), thể hiện vai trò của quá trình truyền dẫn ngoại dịch này chính là kiểm soát các ion và các chất dẫn truyền thần kinh nhằm mục đích giữ cân bằng nội môi.[26] Có xấp xỉ khoảng 78% nơron NGF mà các cúc tận cùng của nó không vận hành tạo synap như ở các nơron điển hình khác. Đây cũng đồng thời là bằng chứng đầu tiên cũng như xác đáng nhất về cơ chế truyền tin hóa học hiện diện ngay cả khi các synap chúng không tồn tại.[25]
Synap điện
[sửa | sửa mã nguồn]Synap điện là synap được hình thành bởi liên kết khe (gap junction) giữa nơron trước synap và nơron sau synap của hai nơron tiếp giáp nhau, cho phép sự dẫn truyền xung động điện thế một cách trực tiếp giữa chúng. Tại synap điện, khoảng cách giữa các nơron chỉ chiếm khoảng 3.5 nm, trong khi đó ở synap hóa các nơron cách nhau xa hơn nhiều khoảng từ 20 đến 40 nm.[27][28] Trái ngược với lại synap hóa học, điện thế sau synap ở synap điện được khởi động không phải là kết quả của việc mở các kênh ion dưới các tác động của chất dẫn truyền thần kinh, nói một cách chính xác hơn do bởi xung động thần kinh lan truyền trực tiếp giữa hai nơron. Đặc biệt synap điện dẫn truyền nhanh hơn nhiều so với synap hóa.[13] Synap điện được thấy ở toàn bộ và xuyên suốt cả hệ thống thần kinh, gồm có võng mạc, nhân lưới trong đồi thị, lớp vỏ não mới, và ở hồi hải mã.[29] Khi mà synap hóa học được tìm thấy ở cả nơron kích thích và nơron ức chế, thì synap điện hầu hết chúng lại hiện diện ở các nơron ức chế cục bộ với kích thước nhỏ. Các synap điện có thể hình thành giữa hai sợi trục, hai sợi nhánh, hoặc giữa sợi trục với sợi nhánh.[30][31] Ở nhiều loài cá và lưỡng cư, các synap điện có thể được thấy ở các cúc tận cùng của synap hóa học, điển hình là nằm trong các tế bào Mauthner.[32]
Những ảnh hưởng do thuốc gây ra
[sửa | sửa mã nguồn]Một trong những đặc tính quan trọng nhất của synap hóa học chính là nằm ở chỗ nó là vị trí nhắm đến của đa số các loại thuốc tâm thần tác động đến trí tuệ và hành vi. Synap chịu tác động bởi các thứ thuốc hay các chất chẳng hạn như curare, strychnine, cocaine, morphine, alcohol, LSD, và còn nhiều hơn thế nữa. Những loại thuốc này chúng tác dụng lên chức năng của synap theo những cách khác nhau, và thường là chỉ xoay quanh các chất dẫn truyền thần kinh đặc hiệu. Điển hình như là, chất curare gây độc bằng cơ chế chặn đứng phân tử truyền đạt acetylcholine làm cho quá trình khử cực ở màng sau synap không thể diễn ra, cuối cùng dẫn đến tê liệt. Strychnine thì ngăn tác dụng ức chế của chất dẫn truyền thần kinh glycine, được biết với tác dụng là bỏ qua các kích thích và làm giảm trương lực cơ toàn cơ thể, kìm hãm ức chế kết quả trương lực cơ tăng mạnh dẫn đến co cơ không kiểm soát (muscle spasms). Morphine tác động ở các synap sử dụng chất truyền đạt endorphin, và chất alcohol tức cồn rượu tăng cường hiệu quả ức chế của chất dẫn truyền thần kinh GABA. Thuốc LSD cản trở các synap vận hành serotonin. Chất cocaine không cho quá trình tái hấp thu dopamine diễn ra và vì thế làm hoạt hóa hệ thống thưởng lâu hơn.
Lịch sử và từ nguyên
[sửa | sửa mã nguồn]Trong những năm 1950, Bernard Katz và Paul Fatt quan sát các dòng điện tự sinh tại synap thần kinh - cơ ở con ếch. Dựa trên những gì đã quan sát được, họ đã phát triển 'giả thuyết lượng tử' chính là nền tảng cơ sở nhằm giải thích được dòng điện sinh học sinh ra bởi do sự giải phóng các chất dẫn truyền thần kinh thông qua cơ chế xuất bào. Chính vì công trình nghiên cứu mang tính đột phá đó mà ông (Katz) đã vinh dự nhận giải thưởng Nobel sinh lý học y khoa vào năm 1970. Vào cuối những năm 1960, Ricardo Miledi và Katz cùng nhau phát triển giả thuyết sự lưu chuyển của dòng ion calci nội bào làm thúc đẩy quá trình xuất bào (giải phóng các phân tử truyền đạt) và gây ra hiện tượng khử cực.
Ngài Charles Scott Sherringtonin đặt ra thuật ngữ 'synap' và lịch sử của từ ngữ này được thể hiện thông qua bức thư mà Sherrington viết gửi cho John Fulton:
'Tôi cảm thấy sự cần thiết lúc này là đưa ra một cái tên nào đó để gọi nhằm ám chỉ đến đó là nơi kết nối giữa tế bào thần kinh với tế bào thần kinh... Tôi thấy nên dùng từ "syndesm" ("dây chằng")... Thầy của tôi [ Ngài Michael Foster ] đã thảo luận với người bạn cũng cùng làm việc tại đại học Trinity, chính là thầy Verrall, học giả với kiến thức uyên thâm về văn chương Hy Lạp đặc biệt thể loại kịch, và thầy Verrall đã nêu ra từ "synap" (bắt nguồn từ Hy Lạp nghĩa là "cái móc").'–Charles Scott Sherringtonin[4]
Xem thêm
[sửa | sửa mã nguồn]- Tính thích nghi của nơron
- Khoa học thần kinh
- Dải synap
Tham khảo
[sửa | sửa mã nguồn]- ^ Drachman D (2005). "Do we have brain to spare?". Neurology. 64 (12): 2004–5. doi:10.1212/01.WNL.0000166914.38327.BB. PMID 15985565.
- ^ Alonso-Nanclares L, Gonzalez-Soriano J, Rodriguez JR, DeFelipe J (tháng 9 năm 2008). "Gender differences in human cortical synaptic density". Proc. Natl. Acad. Sci. U.S.A. 105 (38): 14615–9. Bibcode:2008PNAS..10514615A. doi:10.1073/pnas.0803652105. PMC 2567215. PMID 18779570.{{Chú thích web}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Brain Facts and Figures Washington University.
- ^ a b Cowan, W. Maxwell; Südhof, Thomas C.; Stevens, Charles F. (2003). Synapses. JHU Press. tr. 11. Truy cập ngày 9 tháng 6 năm 2020.
- ^ Rapport, Richard L. (2005). Nerve Endings: The Discovery of the Synapse. W. W. Norton & Company. tr. 1–37. ISBN 978-0-393-06019-5.
- ^ Squire, Larry R.; Floyd Bloom; Nicholas Spitzer (2008). Fundamental Neuroscience. Academic Press. tr. 425–6. ISBN 978-0-12-374019-9.
- ^ Hyman, Steven E.; Eric Jonathan Nestler (1993). The Molecular Foundations of Psychiatry. American Psychiatric Pub. tr. 425–6. ISBN 978-0-88048-353-7.
- ^ Smilkstein, Rita (2003). We're Born to Learn: Using the Brain's Natural Learning Process to Create Today's Curriculum. Corwin Press. tr. 56. ISBN 978-0-7619-4642-7.
- ^ a b Lytton, William W. (2002). From Computer to Brain: Foundations of Computational Neuroscience. Springer. tr. 28. ISBN 978-0-387-95526-1. Axons connecting dendrite to dendrite are dendrodendritic synapses. Axons which connect axon to dendrite are called axodendritic synapses
- ^ a b Garber, Steven D. (2002). Biology: A Self-Teaching Guide. John Wiley and Sons. tr. 175. ISBN 978-0-471-22330-6. synapses connect axons to cell body.
- ^ a b Weiss, Mirin; Dr Steven M. Mirin; Dr Roxanne Bartel (1994). Cocaine. American Psychiatric Pub. tr. 52. ISBN 978-1-58562-138-5. Truy cập ngày 26 tháng 12 năm 2008. Axons terminating on the postsynaptic cell body are axosomatic synapses. Axons that terminate on axons are axoaxonic synapses
- ^ a b c d Widrow, Bernard; Kim, Youngsik; Park, Dookun; Perin, Jose Krause (2019). "Nature's Learning Rule". Artificial Intelligence in the Age of Neural Networks and Brain Computing. Elsevier. tr. 1–30. doi:10.1016/b978-0-12-815480-9.00001-3. ISBN 978-0-12-815480-9.
- ^ a b Kandel 2000, tr. 182
- ^ Bear, Conners, Paradiso (2007). Neuroscience: exploring the brain. Philadelphia, PA: Lippincott Williams & Wilkins. tr. 113–118.{{Chú thích sách}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ Llinás R, Steinberg IZ, Walton K (1981). "Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse". Biophysical Journal. Quyển 33 số 3. tr. 323–351. Bibcode:1981BpJ....33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850.[liên kết hỏng]
- ^ Chapman, Edwin R. (2002). "Synaptotagmin: A Ca2+ sensor that triggers exocytosis?". Nature Reviews Molecular Cell Biology (bằng tiếng Anh). Quyển 3 số 7. tr. 498–508. doi:10.1038/nrm855. ISSN 1471-0080. PMID 12094216.
- ^ Craig C. Garner and Kang Shen. Structure and Function of Vertebrate and Invertebrate Active Zones. Structure and Functional Organization of the Synapse. Ed: Johannes Hell and Michael Ehlers. Springer, 2008.
- ^ Romanov, Roman A.; Lasher, Robert S.; High, Brigit; Savidge, Logan E.; Lawson, Adam; Rogachevskaja, Olga A.; Zhao, Haitian; Rogachevsky, Vadim V.; Bystrova, Marina F.; Churbanov, Gleb D.; Adameyko, Igor; Harkany, Tibor; Yang, Ruibiao; Kidd, Grahame J.; Marambaud, Philippe; Kinnamon, John C.; Kolesnikov, Stanislav S.; Finger, Thomas E. (2018). "Chemical synapses without synaptic vesicles: Purinergic neurotransmission through a CALHM1 channel-mitochondrial signaling complex". Science Signaling. Quyển 11 số 529. tr. eaao1815. doi:10.1126/scisignal.aao1815. ISSN 1945-0877. PMC 5966022. PMID 29739879.
- ^ a b Sherwood L., stikawy (2007). Human Physiology 6e: From Cells to Systems
- ^ Díaz-Ríos M, Miller MW (tháng 6 năm 2006). "Target-specific regulation of synaptic efficacy in the feeding central pattern generator of Aplysia: potential substrates for behavioral plasticity?". Biol. Bull. Quyển 210 số 3. tr. 215–29. doi:10.2307/4134559. JSTOR 4134559. PMID 16801496.
- ^ a b Rang, H.P.; Dale, M.M.; Ritter, J.M. (2003). Pharmacology (ấn bản thứ 5). Edinburgh: Churchill Livingstone. tr. 129. ISBN 978-0-443-07145-4.
- ^ Bruce Alberts; Alexander Johnson; Julian Lewis; Martin Raff; Keith Roberts; Peter Walter, biên tập (2002). "Ch. 11. Section: Single Neurons Are Complex Computation Devices". Molecular Biology of the Cell (ấn bản thứ 4). Garland Science. ISBN 978-0-8153-3218-3.
- ^ Zoli M, Torri C, Ferrari R, và đồng nghiệp (1998). "The emergence of the volume transmission concept". Brain Res. Brain Res. Rev. Quyển 26 số 2–3. tr. 136–47. doi:10.1016/S0165-0173(97)00048-9. PMID 9651506.
- ^ Fuxe K, Dahlström A, Höistad M, và đồng nghiệp (2007). "From the Golgi-Cajal mapping to the transmitter-based characterization of the neuronal networks leading to two modes of brain communication: wiring and volume transmission" (PDF). Brain Res Rev. Quyển 55 số 1. tr. 17–54. doi:10.1016/j.brainresrev.2007.02.009. PMID 17433836.
- ^ a b Oláh S, Füle M, Komlósi G, và đồng nghiệp (2009). "Regulation of cortical microcircuits by unitary GABA-mediated volume transmission". Nature. Quyển 461 số 7268. tr. 1278–81. Bibcode:2009Natur.461.1278O. doi:10.1038/nature08503. PMC 2771344. PMID 19865171.
- ^ Rózsa M, Baka J, Bordé S, Rózsa B, Katona G, Tamás G, và đồng nghiệp (2015). "Unitary GABAergic volume transmission from individual interneurons to astrocytes in the cerebral cortex" (PDF). Brain Structure and Function. Quyển 222 số 1. tr. 651–659. doi:10.1007/s00429-015-1166-9. PMID 26683686.
- ^ Kandel 2000, tr. 176
- ^ Hormuzdi 2004
- ^ Connors BW, Long MA (2004). "Electrical synapses in the mammalian brain". Annu. Rev. Neurosci. Quyển 27 số 1. tr. 393–418. doi:10.1146/annurev.neuro.26.041002.131128. PMID 15217338.
- ^ Veruki ML, Hartveit E (tháng 12 năm 2002). "Electrical synapses mediate signal transmission in the rod pathway of the mammalian retina". J. Neurosci. Quyển 22 số 24. tr. 10558–66. doi:10.1523/JNEUROSCI.22-24-10558.2002. PMC 6758447. PMID 12486148.
- ^ Bennett MV, Pappas GD, Aljure E, Nakajima Y (tháng 3 năm 1967). "Physiology and ultrastructure of electrotonic junctions. II. Spinal and medullary electromotor nuclei in mormyrid fish". J. Neurophysiol. Quyển 30 số 2. tr. 180–208. doi:10.1152/jn.1967.30.2.180. PMID 4167209.
- ^ Pereda AE, Rash JE, Nagy JI, Bennett MV (tháng 12 năm 2004). "Dynamics of electrical transmission at club endings on the Mauthner cells". Brain Res. Brain Res. Rev. Quyển 47 số 1–3. tr. 227–44. doi:10.1016/j.brainresrev.2004.06.010. PMID 15572174.
Đọc thêm
[sửa | sửa mã nguồn]- Carlson, Neil R. (2007). Physiology of Behavior (ấn bản thứ 9). Boston, MA: Pearson Education. ISBN 978-0-205-59389-7.
- Kandel, Eric R.; Schwartz, James H.; Jessell, Thomas M. (2000). Principles of Neural Science (ấn bản thứ 4). New York: McGraw-Hill. ISBN 978-0-8385-7701-1.
- Llinás R, Sugimori M, Simon SM (tháng 4 năm 1982). "Transmission by presynaptic spike-like depolarization in the squid giant synapse". Proc. Natl. Acad. Sci. U.S.A. Quyển 79 số 7. tr. 2415–9. Bibcode:1982PNAS...79.2415L. doi:10.1073/pnas.79.7.2415. PMC 346205. PMID 6954549.
- Llinás R, Steinberg IZ, Walton K (1981). "Relationship between presynaptic calcium current and postsynaptic potential in squid giant synapse". Biophysical Journal. Quyển 33 số 3. tr. 323–352. Bibcode:1981BpJ....33..323L. doi:10.1016/S0006-3495(81)84899-0. PMC 1327434. PMID 6261850.
- Bear, Mark F.; Connors, Barry W.; Paradiso, Michael A. (2001). Neuroscience: Exploring the Brain. Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-3944-3.
- Hormuzdi, SG; Filippov, MA; Mitropoulou, G; Monyer, H; Bruzzone, R (tháng 3 năm 2004). "Electrical synapses: a dynamic signaling system that shapes the activity of neuronal networks". Biochim Biophys Acta. Quyển 1662 số 1–2. tr. 113–137. doi:10.1016/j.bbamem.2003.10.023. PMID 15033583.
- Karp, Gerald (2005). Cell and Molecular Biology: concepts and experiments (ấn bản thứ 4). Hoboken, NJ: John Wiley & Sons. ISBN 978-0-471-46580-5.
- Nicholls, J.G.; Martin, A.R.; Wallace, B.G.; Fuchs, P.A. (2001). From Neuron to Brain (ấn bản thứ 4). Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-439-3.
Liên kết ngoài
[sửa | sửa mã nguồn]- Synapse Review for Kids
- Synapses Lưu trữ ngày 13 tháng 9 năm 2007 tại Wayback Machine Biologymad.com (2004)
- Synapse – Cell Centered Database
- Atlas of Ultrastructure Neurocytology A great electron microscope picture gallery assembled by Kristen Harris' lab of synapses and other neuronal structures.
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HTKTƯ |
|   | |||||||||||||
| HTKNV |
| ||||||||||||||
| Neuron/Dây thần kinh |
| ||||||||||||||
| Phần cuối |
| ||||||||||||||
Từ khóa » Thực Phẩm Chức Năng Synap
-
CHỨC NĂNG XINAP - SlideShare
-
Chất Dẫn Truyền Thần Kinh Là Gì? | Vinmec
-
Sự Dẫn Truyền Thần Kinh - Phiên Bản Dành Cho Chuyên Gia
-
Bệnh Parkinson - Rối Loạn Thần Kinh - Phiên Bản Dành Cho Chuyên Gia
-
DHA & GA - “Bộ đôi Vàng” Phát Triển Trí Não Bé | Anmum Vietnam
-
Sinh Lý Thần Kinh - Học Kì II- Năm Học 2018-2019 - Lã Thị Luyến
-
SGS-742 Là Gì? Công Dụng, Dược Lực Học Và Tương Tác Thuốc
-
Các Nhà Thần Kinh Học Tiết Lộ Việc Tăng Sự Kết Nối Giữa Các Tế Bào Não
-
Tế Bào Thần Kinh: Cấu Tạo, Hoạt động Và Chức Năng - YouMed
-
AQAFFRON - Hỗ Trợ Tĩnh Tâm, Ngon Giấc, Minh Thần - Combo 2 Hộp ...
-
Dinh Dưỡng Vàng Cho Phát Triển Tối ưu Não Bộ - Vinamilk
-
Thuốc Nào Hỗ Trợ Cho Học Trò Mùa Thi?