Lipid – Wikipedia Tiếng Việt
Có thể bạn quan tâm

Trong sinh học và hóa sinh, lipid là một phân tử sinh học hòa tan trong dung môi không phân cực.[2] Các dung môi không phân cực thường là hydrocarbon được sử dụng để hòa tan các phân tử lipid hydrocarbon tự nhiên khác không (hoặc không dễ dàng) hòa tan trong nước, bao gồm acid béo, sáp, sterol, vitamin tan trong chất béo (như vitamin A, D, E và K), monoglyceride, diglyceride, triglyceride và phospholipids.
Các chức năng của lipid bao gồm lưu trữ năng lượng, tạo tín hiệu và hoạt động như các thành phần cấu trúc của màng tế bào.[3][4] Lipid có ứng dụng trong ngành công nghiệp mỹ phẩm và thực phẩm cũng như trong công nghệ nano.[5]
Các nhà khoa học đôi khi xác định lipid như các phân tử nhỏ kỵ nước hoặc lưỡng phân; bản chất lưỡng tính của một số lipid cho phép chúng hình thành các cấu trúc như túi, liposome multilamellar / unilamellar hoặc màng trong môi trường nước. Lipid sinh học có nguồn gốc hoàn toàn hoặc một phần từ hai loại tiểu đơn vị sinh hóa riêng biệt hoặc "khối xây dựng": nhóm ketoacyl và isopren.[3] Sử dụng phương pháp này, lipid có thể được chia thành tám loại: axit béo, glycerolipids, glycerophospholipids, sphingolipids, saccharolipids và polyketide (có nguồn gốc từ ngưng tụ các tiểu đơn vị ketoacyl); và lipid sterol và lipid prenol (có nguồn gốc từ sự ngưng tụ của các tiểu đơn vị isopren).
Mặc dù thuật ngữ "lipid" đôi khi được sử dụng như một từ đồng nghĩa với chất béo, chất béo là một nhóm nhỏ của lipid được gọi là triglyceride. Lipid cũng bao gồm các phân tử như axit béo và các dẫn xuất của chúng (bao gồm tri-, di-, monoglyceride và phospholipids), cũng như các chất chuyển hóa có chứa sterol khác như cholesterol.[6] Mặc dù con người và các động vật có vú khác sử dụng các con đường sinh tổng hợp khác nhau để phá vỡ và tổng hợp lipid, một số lipid thiết yếu không thể được thực hiện theo cách này và phải được lấy từ chế độ ăn uống.
Lịch sử
[sửa | sửa mã nguồn]Lipid có thể được coi là các chất hữu cơ tương đối không hòa tan trong nước, hòa tan trong dung môi hữu cơ (rượu, ether, v.v.) thực sự hoặc có khả năng liên quan đến axit béo và được sử dụng bởi các tế bào sống.
Năm 1815, Henri Braconnot phân loại lipid (graisses) theo hai loại, suifs (mỡ rắn hoặc mỡ động vật) và huiles (dầu lỏng).[7] Năm 1823, Michel Eugène Chevreul đã phát triển một phân loại chi tiết hơn, bao gồm dầu, mỡ, mỡ động vật, sáp, nhựa, balsams và dầu dễ bay hơi (hoặc tinh dầu).[8][9][10]
Sự tổng hợp thành công đầu tiên của một phân tử chất béo trung tính là nhờ nhà hóa học Théophile-Jules Pelouze vào năm 1844, khi ông sản xuất Tributyrin bằng cách phản ứng với axit butyric với glycerin với sự hiện diện của axit sulfuric đậm đặc.[11] Vài năm sau, Marcellin Berthelot, một trong những học sinh của Pelouze, đã tổng hợp tristearin và tripalmitin bằng phản ứng của các axit béo tương tự với glycerin khi có mặt hydro chloride ở nhiệt độ cao.[12]
Năm 1827, William Prout nhận ra chất béo ("chất nhờn"), cùng với protein ("album") và carbohydrate ("sacarine"), như một chất dinh dưỡng quan trọng cho người và động vật.[13][14]
Trong một thế kỷ, các nhà hóa học coi "chất béo" chỉ là chất béo đơn giản được tạo thành từ axit béo và glycerol (glyceride), nhưng các dạng mới đã được mô tả sau đó. Theodore Gobley (1847) đã phát hiện ra phospholipid trong não động vật có vú và trứng gà, được ông gọi là " lecithin ". Thudichum được phát hiện trong não người một số phospholipids (cephalin), glycolipids (cerebroside) và sphingolipids (sphingomyelin).[9]
Các thuật ngữ lipoid, lipin, lipide và lipid đã được sử dụng với ý nghĩa khác nhau từ tác giả này đến tác giả khác.[15] Năm 1912, Rosenbloom và Gies đã đề xuất thay thế "lipoid" bằng "lipin".[16] Năm 1920, Bloor đã giới thiệu một phân loại mới cho "lipoids": lipoids đơn giản (mỡ và sáp), lipoids hợp chất (phospholipoids và glycolipoids) và lipoids có nguồn gốc (axit béo, rượu, sterol).[17][18]
Từ "lipide", bắt nguồn từ nguyên từ lipos Hy Lạp (chất béo), được giới thiệu vào năm 1923 bởi dược sĩ người Pháp Gabriel Bertrand.[19] Bertrands bao gồm trong khái niệm không chỉ các chất béo truyền thống (glyceride), mà còn cả "lipoids", với một hiến pháp phức tạp.[9] Mặc dù từ "lipide" đã được nhất trí thông qua bởi Ủy ban quốc tế của Société de Chimie Biologique trong phiên họp toàn thể trên 3 tháng 7 năm 1923. Từ "lipide" sau đó đã bị biến thành "lipid" vì cách phát âm của nó ('lɪpɪd). Trong ngôn ngữ tiếng Pháp, hậu tố "-ide", từ tiếng Hy Lạp cổ đại "-ίδης" (có nghĩa là 'con trai của' hoặc 'hậu duệ của'), luôn luôn được phát âm (ɪd).
Năm 1947, TP Hilditch đã chia lipid thành "lipid đơn giản", với mỡ và sáp (sáp thật, sterol, rượu).
Phân loại
[sửa | sửa mã nguồn]Lipid đã được phân loại thành tám loại bởi tập đoàn Lipid MAPS như sau:
Axit béo
[sửa | sửa mã nguồn]
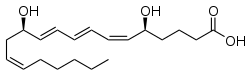
Các axit béo, hoặc dư lượng axit béo khi chúng là một phần của lipid, là một nhóm các phân tử đa dạng được tổng hợp bằng cách kéo dài chuỗi của một mồi acetyl-CoA với các nhóm malonyl-CoA hoặc methylmalonyl-CoA trong một quá trình gọi là tổng hợp axit béo. Chúng được làm bằng chuỗi hydrocarbon kết thúc với nhóm axit cacboxylic; sự sắp xếp này làm cho phân tử có đầu cực, ưa nước và đầu không cực, kỵ nước không tan trong nước. Cấu trúc axit béo là một trong những loại lipid sinh học cơ bản nhất và thường được sử dụng như một khối xây dựng của các lipid phức tạp hơn về cấu trúc. Chuỗi carbon, thường dài từ bốn đến 24 carbons, có thể bão hòa hoặc không bão hòa, và có thể được gắn vào các nhóm chức có chứa oxy, halogen, nitơ và lưu huỳnh. Nếu một axit béo chứa liên kết đôi, có khả năng là đồng phân hình học cis hoặc trans, ảnh hưởng đáng kể đến cấu hình của phân tử. CIS trái phiếu Double nguyên nhân chuỗi axit béo để uốn cong, một hiệu ứng mà trở nên phức tạp với liên kết đôi hơn trong chuỗi. Ba liên kết đôi trong axit linolenic 18 carbon, chuỗi acyl chất béo dồi dào nhất của màng thylakoid thực vật, làm cho các màng này có tính lỏng cao bất chấp nhiệt độ thấp của môi trường, và cũng làm cho axit linolenic vượt trội ở độ phân giải cao ở độ phân giải cao 13- C phổ NMR của lục lạp. Điều này lần lượt đóng một vai trò quan trọng trong cấu trúc và chức năng của màng tế bào.[20] Hầu hết các axit béo tự nhiên là của cấu hình cis, mặc dù dạng trans không tồn tại trong một số chất béo và dầu tự nhiên và một phần hydro hóa.
Ví dụ về các axit béo quan trọng về mặt sinh học bao gồm eicosanoids, có nguồn gốc chủ yếu từ axit arachidonic và axit eicosapentaenoic, bao gồm các loại tiền chất, leukotrien và thromboxan. Axit docosahexaenoic cũng rất quan trọng trong các hệ thống sinh học, đặc biệt là về thị giác.[21][22] Các lớp lipid chính khác trong loại axit béo là các este béo và amit béo. Các este béo bao gồm các chất trung gian sinh hóa quan trọng như este sáp, coenzyme thioester axit béo A, các dẫn xuất ACP của axit béo và các axit béo Carnitine. Các amit béo bao gồm N-acyl ethanolamines, chẳng hạn như chất dẫn truyền thần kinh cannabinoid anandamide.
Glycerolipid
[sửa | sửa mã nguồn]
Glycerolipids bao gồm các mono-, di-, và tri-glycerols được thay thế, những nổi tiếng nhất là acid béo triesters của glycerol, được gọi là triglycerides. Từ "triacylglycerol" đôi khi được sử dụng đồng nghĩa với "triglyceride". Trong các hợp chất này, ba nhóm hydroxyl của glycerol đều được ester hóa, điển hình là bởi các axit béo khác nhau. Bởi vì chúng hoạt động như một kho dự trữ năng lượng, những lipid này chiếm phần lớn chất béo lưu trữ trong các mô động vật. Quá trình thủy phân các liên kết este của triglyceride và giải phóng glycerol và axit béo từ mô mỡ là những bước đầu tiên trong chuyển hóa chất béo.[23]
Các phân lớp bổ sung của glycerolipids được đại diện bởi glycosylglycerol, được đặc trưng bởi sự hiện diện của một hoặc nhiều dư lượng đường gắn với glycerol thông qua liên kết glycosid. Ví dụ về các cấu trúc trong thể loại này là digalactosyldiacylglycerol được tìm thấy trong màng thực vật và Seminolipid từ các tế bào tinh trùng của động vật có vú.
Glycerophospholipid
[sửa | sửa mã nguồn]
Glycerophospholipids, thường được gọi là phospholipids (mặc dù spneumomyelins cũng được phân loại là phospholipids), có mặt khắp nơi trong tự nhiên và là thành phần chính của lớp tế bào lipid kép,[24] cũng như tham gia vào quá trình chuyển hóa và truyền tín hiệu tế bào. Mô thần kinh (bao gồm não) chứa một lượng glycerophospholipids tương đối cao, và sự thay đổi trong thành phần của chúng đã liên quan đến các rối loạn thần kinh khác nhau.[25] Glycerophospholipids có thể được phân chia thành các nhóm riêng biệt, dựa trên bản chất của nhóm đầu cực ở vị trí sn -3 của xương sống glycerol ở sinh vật nhân chuẩn và eubacteria, hoặc vị trí sn -1 trong trường hợp vi khuẩn cổ.
Ví dụ về glycerophospholipids được tìm thấy trong màng sinh học là phosphatidylcholine (còn được gọi là PC, GPCho hoặc lecithin), phosphatidylethanolamine (PE hoặc GPEtn) và phosphatidylserine (PS hoặc GPSer). Ngoài vai trò là thành phần chính của màng tế bào và vị trí gắn kết của protein nội bào và tế bào, một số glycerophospholipids trong các tế bào nhân chuẩn, như phosphatidylinositols và axit phosphatidic là tiền chất của hoặc chính chúng, là chất dẫn truyền thứ hai của màng.[26] Thông thường, một hoặc cả hai nhóm hydroxyl được acylated với các axit béo chuỗi dài, nhưng cũng có những alkyl-liên kết và 1Z-alkenyl liên kết (plasmalogen) glycerophospholipids, cũng như dialkylether biến thể trong archaebacteria.
Sphingolipid
[sửa | sửa mã nguồn]
Sphingolipids là một họ các hợp chất phức tạp [27] có chung đặc điểm cấu trúc, một xương sống cơ bản được tổng hợp de novo từ serine amino acid và acyl CoA chuỗi dài, sau đó chuyển thành ceramide, phosphosphingolipids và glycosphingolipids khác Các hợp chất. Các cơ sở chính sprialoid của động vật có vú thường được gọi là sprialosine. Ceramides (cơ sở N-acyl-sphingoid) là một phân lớp chính của các dẫn xuất cơ sở spakenoid với một axit béo liên kết amide. Các axit béo thường bão hòa hoặc không bão hòa đơn với độ dài chuỗi từ 16 đến 26 nguyên tử carbon.[28]
Các phosphosphingolipids chính của động vật có vú là sprialomyelins (ceramide phosphocholines), trong khi côn trùng chứa chủ yếu là ceramide phosphoethanolamines và nấm có phytoceramide phosphoinositols và nhóm đầu chứa mannose. Các glycosphingolipids là một họ các phân tử đa dạng bao gồm một hoặc nhiều dư lượng đường được liên kết thông qua một liên kết glycosid với cơ sở spakenoid. Ví dụ trong số này là các glycosphingolipids đơn giản và phức tạp như cerebroside và ganglioside.
Sterol
[sửa | sửa mã nguồn]
Sterol, chẳng hạn như cholesterol và các dẫn xuất của nó, là một thành phần quan trọng của lipid màng, cùng với glycerophospholipids và sprialomyelins. Các ví dụ khác của sterol là axit mật và liên hợp của chúng, trong động vật có vú là dẫn xuất oxy hóa của cholesterol và được tổng hợp ở gan. Tương đương thực vật là các phytosterol, như -sitosterol, stigmasterol và Brassicasterol; hợp chất thứ hai cũng được sử dụng như một dấu ấn sinh học cho sự phát triển của tảo. Sterol chiếm ưu thế trong màng tế bào nấm là ergosterol.
Sterol là steroid trong đó một trong những nguyên tử hydro được thay thế bằng nhóm hydroxyl, ở vị trí 3 trong chuỗi carbon. Chúng có điểm chung với các cấu trúc lõi bốn vòng hợp nhất. Steroid có vai trò sinh học khác nhau như hormone và các phân tử tín hiệu. Các steroid mười tám carbon (C18) bao gồm họ estrogen trong khi các steroid C19 bao gồm các androgen như testosterone và androsterone. Phân lớp C21 bao gồm proestogen cũng như glucocorticoids và mineralocorticoids.[29] Các secosteroids, bao gồm các dạng vitamin D khác nhau, được đặc trưng bởi sự phân tách của vòng B của cấu trúc lõi.
Prenol
[sửa | sửa mã nguồn]
Các lipid prenol được tổng hợp từ các tiền chất đơn vị năm carbon là isopentenyl diphosphate và dimethylallyl diphosphate được sản xuất chủ yếu thông qua con đường axit mevalonic (MVA). Các isoprenoid đơn giản (rượu tuyến tính, diphosphate, v.v.) được hình thành bằng cách bổ sung liên tiếp các đơn vị C5 và được phân loại theo số lượng các đơn vị terpene này. Các cấu trúc chứa lớn hơn 40 nguyên tử được gọi là polyterpen. Carotenoids là các isoprenoid đơn giản quan trọng có chức năng như chất chống oxy hóa và là tiền chất của vitamin A. Một loại phân tử quan trọng về mặt sinh học khác được minh họa bằng quinon và hydroquinone, có chứa đuôi isoprenoid gắn với lõi quinonoid có nguồn gốc không phải isoprenoid. Vitamin E và vitamin K, cũng như ubiquinones, là những ví dụ của lớp này. Prokaryote tổng hợp polyprenol (được gọi là bactoprenols) trong đó đơn vị isoprenoid cuối cùng gắn với oxy vẫn chưa bão hòa, trong khi ở polyprenol động vật (dolichol) thì isoprenoid cuối bị giảm.
Saccharolipid
[sửa | sửa mã nguồn]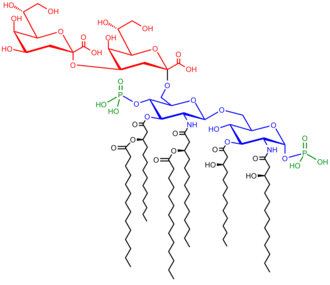
Saccharolipids mô tả các hợp chất trong đó axit béo được liên kết trực tiếp với đường trục, tạo thành các cấu trúc tương thích với màng kép. Trong saccharolipids, một monosacarit thay thế cho xương sống glycerol có trong glycerolipids và glycerophospholipids. Các sacarolipid quen thuộc nhất là tiền chất glucosamine bị acyl hóa của Lipid Một thành phần của lipopolysacarit ở vi khuẩn gram âm. Lipid điển hình Một phân tử là disacarit của glucosamine, được tạo dẫn xuất với tới bảy chuỗi acyl béo. Lipopolysacarit tối thiểu cần cho sự tăng trưởng của E. coli là Kdo 2 -Lipid A, một disacarit hexa-acylated của glucosamine được glycosyl hóa với hai dư lượng axit 3-deoxy-D-manno-octulonic (Kdo).
Polyketide
[sửa | sửa mã nguồn]Polyketide được tổng hợp bằng cách trùng hợp các tiểu đơn vị acetyl và propionyl bằng các enzyme cổ điển cũng như các enzyme lặp và đa bào có chung đặc điểm cơ học với các chất tổng hợp axit béo. Chúng bao gồm nhiều chất chuyển hóa thứ cấp và các sản phẩm tự nhiên từ các nguồn động vật, thực vật, vi khuẩn, nấm và biển, và có sự đa dạng về cấu trúc lớn. Nhiều polyketide là các phân tử tuần hoàn có xương sống thường được sửa đổi thêm bằng cách glycosyl hóa, methyl hóa, hydroxyl hóa, oxy hóa hoặc các quá trình khác. Nhiều chất chống vi khuẩn, chống ký sinh trùng và chống ung thư thường được sử dụng là các dẫn xuất polyketide hoặc polyketide, như erythromycins, tetracycline, avermectin và epothilone chống ung thư.
Chức năng sinh học
[sửa | sửa mã nguồn]Màng lọc
[sửa | sửa mã nguồn]Các tế bào nhân chuẩn có các bào quan liên kết màng ngăn cách thực hiện các chức năng sinh học khác nhau. Các glycerophospholipids là thành phần cấu trúc chính của màng sinh học, như màng sinh chất của tế bào và màng nội bào của bào quan; trong tế bào động vật, màng sinh chất ngăn cách các thành phần nội bào với môi trường ngoại bào. Các glycerophospholipids là amphipathic phân tử (có chứa cả kỵ nước và ưa nước vùng) có chứa một lõi glycerol liên kết với hai béo "đuôi" có nguồn gốc từ axit bằng cách este mối liên kết và một nhóm "đầu" bởi phosphate liên kết este. Trong khi glycerophospholipids là thành phần chính của màng sinh học, các thành phần lipid không glyceride khác như sphingomyelin và sterol (chủ yếu là cholesterol trong màng tế bào động vật) cũng được tìm thấy trong màng sinh học.[30] Trong thực vật và tảo, galactosyldiacylglycerol,[31] và sulfoquinovosyldiacylglycerol, không có nhóm phosphat, là thành phần quan trọng của màng của lục lạp và các bào quan có liên quan và là các lipid có nhiều nhất trong các mô quang hợp và một số vi khuẩn.
Màng thylakoid thực vật có thành phần lipid lớn nhất của một loại monogalactosyl diglyceride (MGDG) không hai lớp, và ít phospholipid; Mặc dù có thành phần lipid độc đáo này, màng thylakoid lục lạp đã được chứng minh có chứa ma trận hai lớp lipid động như được tiết lộ bởi các nghiên cứu cộng hưởng từ và kính hiển vi điện tử.
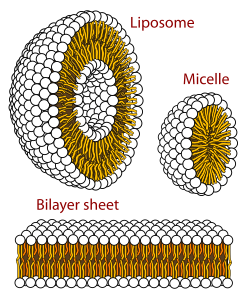
Một màng sinh học là một dạng của hai lớp lipid pha lamellar. Sự hình thành của hai lớp lipid là một quá trình ưa thích năng lượng khi glycerophospholipids được mô tả ở trên trong một môi trường nước.[32] Điều này được gọi là hiệu ứng kỵ nước. Trong một hệ thống nước, các đầu cực của lipid thẳng hàng với môi trường cực, nước, trong khi đuôi kỵ nước giảm thiểu sự tiếp xúc của chúng với nước và có xu hướng tụ lại với nhau, tạo thành một túi; tùy thuộc vào nồng độ của lipid, tương tác sinh lý này có thể dẫn đến sự hình thành các mixen, liposome hoặc hai lớp lipid. Các tập hợp khác cũng được quan sát và tạo thành một phần của tính đa hình của hành vi amphiphile (lipid). Hành vi giai đoạn là một lĩnh vực nghiên cứu trong vật lý sinh học và là chủ đề của hiện tại [khi nào?] nghiên cứu học thuật. Các mixen và hai lớp hình thành trong môi trường cực bởi một quá trình được gọi là hiệu ứng kỵ nước. Khi hòa tan một chất lipophilic hoặc amphiphilic trong môi trường cực, các phân tử cực (tức là nước trong dung dịch nước) trở nên có trật tự hơn xung quanh chất lipophilic hòa tan, vì các phân tử cực không thể tạo liên kết hydro với các vùng lipophilic của amphiphile. Vì vậy, trong một môi trường nước, các phân tử nước tạo thành một cái lồng " clathrate " được sắp xếp xung quanh phân tử lipophilic hòa tan.
Sự hình thành lipid thành màng protocell đại diện cho một bước quan trọng trong các mô hình abiogenesis, nguồn gốc của sự sống.
Lưu trữ năng lượng
[sửa | sửa mã nguồn]Triglyceride, được lưu trữ trong mô mỡ, là một hình thức lưu trữ năng lượng chính cả ở động vật và thực vật. Chúng là một nguồn năng lượng chính vì carbohydrate là cấu trúc giảm hoàn toàn. So với glycogen chỉ đóng góp một nửa năng lượng trên khối lượng tinh khiết của nó, các chất béo trung tính được liên kết với hydrogens, không giống như trong carbohydrate.[33] Các tế bào mỡ, hay tế bào mỡ, được thiết kế để tổng hợp và phân hủy triglyceride liên tục ở động vật, với sự phân hủy chủ yếu bằng cách kích hoạt enzyme lipase nhạy cảm với hormone. Quá trình oxy hóa hoàn toàn các axit béo cung cấp hàm lượng calo cao, khoảng 38 kJ / g (9 kcal / g), so với 17 kJ / g (4 kcal / g) cho sự phân hủy carbohydrate và protein. Những con chim di cư phải bay đường dài mà không ăn, sử dụng năng lượng dự trữ của chất béo trung tính để cung cấp nhiên liệu cho các chuyến bay của chúng.[34]
Báo hiệu
[sửa | sửa mã nguồn]Trong những năm gần đây, bằng chứng đã xuất hiện cho thấy tín hiệu lipid là một phần quan trọng của tín hiệu tế bào.[35][36] Tín hiệu lipid có thể xảy ra thông qua kích hoạt các thụ thể hạt nhân hoặc protein G, và các thành viên của một số loại lipid khác nhau đã được xác định là các phân tử tín hiệu và sứ giả tế bào. Chúng bao gồm sphingosine-1-phosphate, một loại sp Breatholipid có nguồn gốc từ ceramide là một phân tử truyền tin mạnh có liên quan đến việc điều chỉnh huy động calci, tăng trưởng tế bào và apoptosis; diacylglycerol (DAG) và phosphatidylinositol phosphat (PIPs), liên quan đến hoạt hóa qua trung gian calci của protein kinase C; Prostaglandin, một loại eicosanoid có nguồn gốc axit béo liên quan đến viêm và miễn dịch; các hormone steroid như estrogen, testosterone và cortisol, điều chỉnh một loạt các chức năng như sinh sản, trao đổi chất và huyết áp; và các ox-quang như 25-hydroxy-cholesterol là chất chủ vận thụ thể gan X. Lipid Phosphatidylserine được biết là có liên quan đến việc truyền tín hiệu cho quá trình thực bào của các tế bào apoptotic hoặc các mảnh của tế bào. Họ thực hiện điều này bằng cách tiếp xúc với mặt ngoại bào của màng tế bào sau khi vô hiệu hóa các flippase đặt chúng ở phía tế bào học và kích hoạt scramblase, làm xáo trộn sự định hướng của phospholipids. Sau khi điều này xảy ra, các tế bào khác nhận ra phosphatidylserine và phagocytosize các tế bào hoặc các mảnh tế bào phơi bày chúng.
Các chức năng khác
[sửa | sửa mã nguồn]Các vitamin "tan trong chất béo" (A, D, E và K) - đó là lipid dựa trên isopren - là các chất dinh dưỡng thiết yếu được lưu trữ trong gan và các mô mỡ, với một loạt các chức năng. Acyl-Carnitines tham gia vào quá trình vận chuyển và chuyển hóa axit béo trong và ngoài ty thể, nơi chúng trải qua quá trình oxy hóa beta.[37] Polyprenol và các dẫn xuất phosphoryl hóa của chúng cũng đóng vai trò vận chuyển quan trọng, trong trường hợp này là vận chuyển oligosacarit qua màng. Polyprenol phosphate đường và polyprenol diphosphate đường chức năng trong các phản ứng glycosyl hóa ngoài tế bào chất, trong sinh tổng hợp polysaccharide ngoại bào (ví dụ, peptidoglycan trùng hợp trong vi khuẩn), và trong eukaryotic protein N- glycosyl hóa. Cardiolipin là một phân lớp của glycerophospholipids chứa bốn chuỗi acyl và ba nhóm glycerol đặc biệt có nhiều trong màng ty thể bên trong. Chúng được cho là kích hoạt các enzyme liên quan đến quá trình phosphoryl oxy hóa. Lipid cũng tạo thành cơ sở của hormone steroid.[38]
Sự trao đổi chất
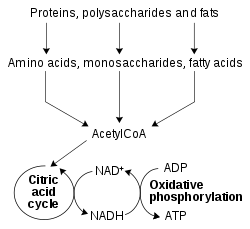
[sửa | sửa mã nguồn]Các chất béo chính trong chế độ ăn uống cho người và các động vật khác là triglyceride động vật và thực vật, sterol và phospholipids màng. Quá trình chuyển hóa lipid tổng hợp và làm suy giảm các cửa hàng lipid và tạo ra các lipid cấu trúc và chức năng đặc trưng của các mô riêng lẻ.
Sinh tổng hợp
[sửa | sửa mã nguồn]Ở động vật, khi có sự cung cấp quá nhiều carbohydrate trong chế độ ăn uống, lượng carbohydrate dư thừa sẽ được chuyển thành triglyceride. Điều này liên quan đến việc tổng hợp axit béo từ acetyl-CoA và quá trình ester hóa axit béo trong sản xuất triglyceride, một quá trình gọi là lipogenesis.[39] Các axit béo được tạo ra bởi các chất tổng hợp axit béo trùng hợp và sau đó làm giảm các đơn vị acetyl-CoA. Các chuỗi acyl trong các axit béo được kéo dài bằng một chu kỳ phản ứng thêm nhóm acetyl, khử nó thành rượu, khử nước thành nhóm anken và sau đó khử lại thành nhóm ankan. Các enzyme của sinh tổng hợp axit béo được chia thành hai nhóm, ở động vật và nấm, tất cả các phản ứng tổng hợp axit béo này được thực hiện bởi một protein đa chức năng duy nhất, trong khi ở plastid thực vật và các enzyme của vi khuẩn thực hiện từng bước trên con đường. Các axit béo sau đó có thể được chuyển đổi thành triglyceride được đóng gói trong lipoprotein và được tiết ra từ gan.
Sự tổng hợp các axit béo không bão hòa liên quan đến phản ứng khử bão hòa, theo đó một liên kết đôi được đưa vào chuỗi acyl béo. Ví dụ, ở người, quá trình khử axit stearic của stearoyl-CoA desaturase-1 tạo ra axit oleic. Axit linoleic axit béo không bão hòa kép cũng như axit α-linolenic không bão hòa ba lần không thể được tổng hợp trong các mô của động vật có vú, và do đó là các axit béo thiết yếu và phải được lấy từ chế độ ăn uống.[40]
Sự tổng hợp triglyceride diễn ra trong mạng lưới nội chất bằng con đường trao đổi chất trong đó các nhóm acyl trong acyl-CoAs béo được chuyển đến các nhóm hydroxyl của glycerol-3-phosphate và diacylglycerol.[41]
Terpenes và isoprenoid, bao gồm các carotenoids, được tạo ra bởi sự lắp ráp và sửa đổi các đơn vị isopren được tặng từ tiền chất phản ứng isopentenyl pyrophosphate và dimethylallyl pyrophosphate. Những tiền chất có thể được thực hiện theo những cách khác nhau. Ở động vật và vi khuẩn cổ, con đường mevalonate tạo ra các hợp chất này từ acetyl-CoA, trong khi ở thực vật và vi khuẩn , con đường không mevalonate sử dụng pyruvate và glyceraldehyd 3-phosphate làm chất nền. Một phản ứng quan trọng sử dụng các nhà tài trợ isoprene kích hoạt này là sinh tổng hợp steroid. Tại đây, các đơn vị isopren được nối với nhau để tạo ra squalene và sau đó gấp lại và tạo thành một bộ vòng để tạo lanosterol. Lanosterol sau đó có thể được chuyển đổi thành các steroid khác như cholesterol và ergosterol.
Phân giải
[sửa | sửa mã nguồn]Quá trình oxy hóa beta là quá trình trao đổi chất nhờ đó các axit béo bị phá vỡ trong ty thể hoặc trong peroxisome để tạo ra acetyl-CoA. Phần lớn, các axit béo bị oxy hóa theo một cơ chế tương tự, nhưng không giống với, một sự đảo ngược của quá trình tổng hợp axit béo. Đó là, các mảnh hai carbon được loại bỏ tuần tự từ đầu carboxyl của axit sau các bước khử hydro, hydrat hóa và oxy hóa để tạo thành axit beta-keto, được phân tách bằng thiolysis. Acetyl-CoA cuối cùng được chuyển đổi thành ATP, CO 2 và H 2 O bằng cách sử dụng chu trình axit citric và chuỗi vận chuyển điện tử. Do đó, chu trình axit citric có thể bắt đầu tại acetyl-CoA khi chất béo bị phân hủy thành năng lượng nếu có ít hoặc không có glucose. Năng lượng năng lượng của quá trình oxy hóa hoàn toàn palmitate axit béo là 106 ATP.[42] Các axit béo không bão hòa và chuỗi lẻ đòi hỏi các bước enzyme bổ sung cho sự phân giải.
Dinh dưỡng và sức khỏe
[sửa | sửa mã nguồn]Hầu hết các chất béo được tìm thấy trong thực phẩm là ở dạng triglyceride, cholesterol và phospholipids. Một số chất béo trong chế độ ăn uống là cần thiết để tạo điều kiện cho việc hấp thụ các vitamin tan trong chất béo (A, D, E và K) và carotenoids.[43] Con người và các động vật có vú khác có yêu cầu về chế độ ăn uống đối với một số axit béo thiết yếu, chẳng hạn như axit linoleic (axit béo omega-6) và axit alpha-linolenic (một loại axit béo omega-3) vì chúng không thể được tổng hợp từ các tiền chất đơn giản trong chế độ ăn uống.[40] Cả hai axit béo này là axit béo không bão hòa đa carbon 18 carbon khác nhau về số lượng và vị trí của các liên kết đôi. Hầu hết các loại dầu thực vật đều giàu axit linoleic (nghệ tây, hướng dương và dầu ngô). Alpha-linolenic acid được tìm thấy trong lá xanh của cây, và trong các loại hạt, hạt và cây họ đậu được chọn lọc (đặc biệt là hạt lanh, hạt cải dầu, quả óc chó và đậu nành). Dầu cá đặc biệt giàu axit béo omega-3 chuỗi dài axit eicosapentaenoic (EPA) và axit docosahexaenoic (DHA).[44] Nhiều nghiên cứu đã chỉ ra những lợi ích sức khỏe tích cực liên quan đến việc tiêu thụ axit béo omega-3 đối với sự phát triển của trẻ sơ sinh, ung thư, bệnh tim mạch và các bệnh tâm thần khác nhau, chẳng hạn như trầm cảm, rối loạn tăng động giảm chú ý và chứng mất trí. Ngược lại, hiện nay người ta đã xác định rõ rằng việc tiêu thụ chất béo chuyển hóa, chẳng hạn như những chất có trong dầu thực vật hydro hóa một phần, là một yếu tố nguy cơ đối với bệnh tim mạch. Chất béo tốt cho bạn có thể biến thành chất béo chuyển hóa bằng cách nấu quá chín.
Một vài nghiên cứu cho thấy rằng tổng lượng chất béo trong chế độ ăn uống có liên quan đến việc tăng nguy cơ béo phì và bệnh tiểu đường. Tuy nhiên, một số nghiên cứu rất lớn, bao gồm Thử nghiệm điều chỉnh chế độ ăn kiêng cho sức khỏe phụ nữ, nghiên cứu kéo dài 8 năm với 49.000 phụ nữ, Nghiên cứu sức khỏe của y tá và nghiên cứu theo dõi chuyên gia y tế, cho thấy không có mối liên hệ nào như vậy. Không có nghiên cứu nào trong số này cho thấy bất kỳ mối liên hệ nào giữa tỷ lệ phần trăm calo từ chất béo và nguy cơ ung thư, bệnh tim hoặc tăng cân. Nguồn dinh dưỡng, một trang web được duy trì bởi Khoa Dinh dưỡng tại Trường Y tế Công cộng Harvard, tóm tắt bằng chứng hiện tại về tác động của chất béo trong chế độ ăn kiêng: "Nghiên cứu chi tiết về nghiên cứu chi tiết được thực hiện tại Harvard cho thấy tổng lượng chất béo trong chế độ ăn uống không thực sự liên quan đến cân nặng hoặc bệnh tật. "
Tham khảo
[sửa | sửa mã nguồn]- ^ Stryer et al., p. 328.
- ^ IUPAC, Compendium of Chemical Terminology (Giản lược thuật ngữ hoá học), bản thứ 2 ("Gold Book") (1997). Bản đã chỉnh sửa trực tuyến: (2006–) "lipids". doi:10.1351/goldbook.L03571
- ^ a b Fahy E, Subramaniam S, Murphy RC, Nishijima M, Raetz CR, Shimizu T, Spener F, van Meer G, Wakelam MJ, Dennis EA (April 2009). "Update of the LIPID MAPS comprehensive classification system for lipids". Journal of Lipid Research. 50 Suppl (S1): S9–14. doi:10.1194/jlr.R800095-JLR200. PMC 2674711. PMID 19098281.
- ^ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes RW, Cotter D, Dinasarapu AR, Maurya MR (October 2011). "Bioinformatics and systems biology of the lipidome". Chemical Reviews. 111 (10): 6452–90. doi:10.1021/cr200295k. PMC 3383319. PMID 21939287.
- ^ Mashaghi S, Jadidi T, Koenderink G, Mashaghi A (February 2013). "Lipid nanotechnology". International Journal of Molecular Sciences. 14 (2): 4242–82. doi:10.3390/ijms14024242. PMC 3588097. PMID 23429269.

- ^ Michelle A, Hopkins J, McLaughlin CW, Johnson S, Warner MQ, LaHart D, Wright JD (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. ISBN [[Đặc_biệt:Nguồn_sách/978-0-13-981176-0|978-0-13-981176-0]] calling template requires template_name parameter.
- ^ Braconnot H (ngày 31 tháng 3 năm 1815). "Sur la nature des corps gras". Annales de chimie. Quyển 2 số XCIII. tr. 225–277.
- ^ Chevreul ME (1823). Recherches sur les corps gras d'origine animale. Paris: Levrault.
- ^ a b c Leray C (2012). Introduction to Lipidomics. Boca Raton: CRC Press. ISBN 9781466551466.
- ^ Leray C (2015). "Introduction, History and Evolution.". Lipids. Nutrition and health. Boca Raton: CRC Press. ISBN 9781482242317.
- ^ Ann Chim Phys 1844, 10, 434
- ^ C R Séances Acad Sci, Paris, 1853, 36, 27; Ann Chim Phys 1854, 41, 216
- ^ "Chronological history of lipid center". Cyberlipid Center. Bản gốc lưu trữ ngày 13 tháng 10 năm 2017. Truy cập ngày 1 tháng 12 năm 2017.
- ^ Prout W (1827). "On the ultimate composition of simple alimentary substances, with some preliminary remarks on the analysis of organised bodies in general". Phil. Trans. tr. 355–388.
- ^ Culling CF (1974). "Lipids. (Fats, Lipoids. Lipins).". Handbook of Histopathological Techniques (ấn bản thứ 3). London: Butterworths. tr. 351–376. ISBN 9781483164793.
- ^ Rosenbloom J, Gies WJ (1911). "Suggestion to teachers of biochemistry. I. A proposed chemical classification of lipins, with a note on the intimate relation between cholesterols and bile salts". Biochem. Bull. Quyển 1. tr. 51–6.
- ^ Bloor WR (1920). "Outline of a classication of the lipids". Proc. Soc. Exp. Biol. Med. Quyển 17 số 6. tr. 138–140. doi:10.3181/00379727-17-75.
- ^ Christie WW, Han X (2010). Lipid Analysis: Isolation, Separation, Identification and Lipidomic Analysis. Bridgwater, England: The Oily Press. ISBN 9780857097866.
- ^ Bertrand G (1923). "Projet de reforme de la nomenclature de Chimie biologique". Bulletin de la Société de Chimie Biologique. Quyển 5. tr. 96–109.
- ^ Devlin, pp. 193–195.
- ^ "A Long Lipid, a Long Name: Docosahexaenoic Acid". The Lipid Chronicles. ngày 2 tháng 12 năm 2011.
- ^ "DHA for Optimal Brain and Visual Functioning". DHA/EPA Omega-3 Institute.
- ^ van Holde and Mathews, pp. 630–31.
- ^ "The Structure of a Membrane". The Lipid Chronicles. ngày 5 tháng 11 năm 2011. Truy cập ngày 31 tháng 12 năm 2011.
- ^ Farooqui AA, Horrocks LA, Farooqui T (tháng 6 năm 2000). "Glycerophospholipids in brain: their metabolism, incorporation into membranes, functions, and involvement in neurological disorders". Chemistry and Physics of Lipids. Quyển 106 số 1. tr. 1–29. doi:10.1016/S0009-3084(00)00128-6. PMID 10878232.
- ^ van Holde and Mathews, p. 844.
- ^ Merrill AH, Sandoff K (2002). "Chapter 14: Sphingolipids: Metabolism and Cell Signaling" (PDF). Trong Vance JE, Vance EE (biên tập). Biochemistry of Lipids, Lipoproteins and Membranes (ấn bản thứ 4). Amsterdam: Elsevier. tr. 373–407. ISBN 978-0-444-51138-6.
- ^ Devlin, pp. 421–422.
- ^ Stryer et al., p. 749.
- ^ Stryer et al., pp. 329–331.
- ^ Heinz E. (1996). "Plant glycolipids: structure, isolation and analysis", pp. 211–332 in Advances in Lipid Methodology, Vol. 3. W.W. Christie (ed.). Oily Press, Dundee. ISBN 978-0-9514171-6-4
- ^ Stryer et al., pp. 333–334.
- ^ Rosen ED, Spiegelman BM (tháng 12 năm 2006). "Adipocytes as regulators of energy balance and glucose homeostasis". Nature. Quyển 444 số 7121. tr. 847–53. doi:10.1038/nature05483. PMC 3212857. PMID 17167472.
- ^ Stryer et al., p. 619.
- ^ Malinauskas T, Aricescu AR, Lu W, Siebold C, Jones EY (tháng 7 năm 2011). "Modular mechanism of Wnt signaling inhibition by Wnt inhibitory factor 1". Nature Structural & Molecular Biology. Quyển 18 số 8. tr. 886–93. doi:10.1038/nsmb.2081. PMC 3430870. PMID 21743455.
- ^ Malinauskas T (tháng 3 năm 2008). "Docking of fatty acids into the WIF domain of the human Wnt inhibitory factor-1". Lipids. Quyển 43 số 3. tr. 227–30. doi:10.1007/s11745-007-3144-3. PMID 18256869.
- ^ Indiveri C, Tonazzi A, Palmieri F (tháng 10 năm 1991). "Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria". Biochimica et Biophysica Acta (BBA) - Biomembranes. Quyển 1069 số 1. tr. 110–6. doi:10.1016/0005-2736(91)90110-t. PMID 1932043.
- ^ "Steroids". Elmhurst. edu. Bản gốc lưu trữ ngày 23 tháng 10 năm 2011. Truy cập ngày 10 tháng 10 năm 2013.
- ^ Stryer et al., p. 634.
- ^ a b Stryer et al., p. 643.
- ^ Stryer et al., pp. 733–739.
- ^ Stryer et al., pp. 625–626.
- ^ Bhagavan, p. 903.
- ^ Bhagavan, p. 388.
Liên kết ngoài
[sửa | sửa mã nguồn]
| |
|---|---|
| Chung |
|
| Hình học |
|
| Axit béo |
|
| Phospholipid |
|
| Steroid |
|
| Sphingolipid |
|
| Eicosanoids |
|
| Glyxerit |
|
| Bản mẫu:Nhóm sinh hóa | |
| |
|---|---|
| Bão hòa |
|
| ω−3 Không bão hòa |
|
| ω−5 Không bão hòa |
|
| ω−6 Không bão hòa |
|
| ω−7 Không bão hòa |
|
| ω−9 Không bão hòa |
|
| ω−10 Không bão hòa |
|
| ω−11 Không bão hòa |
|
| ω−12 Không bão hòa |
|
| |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ceramide |
| ||||||||||
| Đường chuyển hoáGanglioside |
| ||||||||||
| Khác |
| ||||||||||
| |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Thông tin chung |
|    | |||||||||||||||||||||||||||||||
| Chuyển hóa năng lượng |
| ||||||||||||||||||||||||||||||||
| Con đường đặc hiệu |
| ||||||||||||||||||||||||||||||||
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phytochemicals |
| ||||||||||||
| Misc: | List of phytochemicals and foods in which they are prominent | ||||||||||||
| |||||||||||||
| Các họ chính của các chất hóa sinh | |||||||||||||
| Peptit | Amino acid | Axit nucleic | Cacbohydrat | Lipid | Terpen | Carotenoit | Tetrapyrrol | Phụ nhân tử Enzym | Steroit | Flavonoit | Ancaloit | Polyketit | Glicozit | |||||||||||||
| Các chất tương tự của axít nucleic: | Các chất tương tự của axít nucleic: | ||||||||||||
| |
|---|---|
|
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chung |
| ||||||||||||||||||||||||||||||||||
| Công nghiệp thực phẩm |
| ||||||||||||||||||||||||||||||||||
| Chính trị thực phẩm |
| ||||||||||||||||||||||||||||||||||
| Tổ chức |
| ||||||||||||||||||||||||||||||||||
| ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Các thuốcliên quanđến lặn |
|  | ||||||||||||||
| Divingphysiology |
| |||||||||||||||
| Vật lý học của lặn |
| |||||||||||||||
| Môi trường lặn |
| |||||||||||||||
| Các nhànghiên cứu về lặn về mặt y học và sinh lý học |
| |||||||||||||||
| Các tổ chức nghiêncứu y học lặn | Hiệp hội y khoa hàng không Mạng thông báo đa phương tiện (DAN) Trung tâm nghiên cứu bệnh lặn (DDRC) Hội đồng tư vấn y tế lặn (DMAC) Ủy ban công nghệ lặn châu Âu (EDTC) Hội Dưới nước và Bội áp châu Âu (EUBS) Hội đồng Quốc gia về Công nghệ Y học Lặn và Bội áp Phòng thí nghiệm nghiên cứu y học hải quân ngầm Trường Y học Thủy quân Hoàng gia Úc Tổ chức Rubicon Hội Y học dưới nước Nam Thái Bình Dương (SPUMS) Hiệp hội Y khoa dưới nước và Bội áp Nam Phi (SAUHMA) Hiệp hội Y khoa và Bội áp dưới nước (UHMS) Đơn vị lặn thử nghiệm hải quân Hoa Kỳ (NEDU) | |||||||||||||||
| ||||||||||||||||
 Ẩm thực
Ẩm thực Sinh học
Sinh học
| Cơ sở dữ liệu tiêu đề chuẩn | |
|---|---|
| Quốc tế |
|
| Quốc gia |
|
| Khác |
|
Từ khóa » đặc điểm Chung Của Lipid Là Gì
-
Đặc điểm Chung Của Tất Cả Các Loại Lipit Là?
-
Đặc điểm Chung Của Tất Cả Các Loại Lipit Là?
-
[ĐÚNG NHẤT] Đặc điểm Chung Của Các Loại Lipit - TopLoigiai
-
Vai Trò Của Lipid Trong Cơ Thể - Vinmec
-
Nêu đặc Tính Chung, Cấu Trúc , Chức Năng Của Lipit đơn Giản, Lipit ...
-
Lipit Không Có đặc điểm? - Luật Hoàng Phi
-
Đặc điểm Chung Của Tất Cả Các Loại Lipit Là? - Hữu Trí - HOC247
-
Lipid Là Gì? Vai Trò, Cấu Tạo, Tính Chất, Phân Loại
-
Lipit Là Gì? Lý Thuyết, Phân Loại Và Vai Trò Của Lipit - THPT Sóc Trăng
-
Tổng Quan Về Chuyển Hóa Lipid - Phiên Bản Dành Cho Chuyên Gia
-
10 đặc điểm Của Chất Béo Nổi Bật Nhất - Thpanorama
-
Các Loại Lipit Có Chung đặc Tính Là?
-
ID10-301. Đặc điểm Chung Của Tất Cả Các Loại Lipit Là?
-
Lipit Là Gì? Công Thức Cấu Tạo, Chức Năng Và Các Loại Lipit