Rắn – Wikipedia Tiếng Việt
Có thể bạn quan tâm
| Rắn | |
|---|---|
| Thời điểm hóa thạch: Creta muộn – hiện nay,[1] 94–0 triệu năm trước đây TiềnꞒ Ꞓ O S D C P T J K Pg N | |
 Sự đa dạng của rắn Sự đa dạng của rắn | |
| Phân loại khoa học | |
| Giới (regnum) | Animalia |
| Ngành (phylum) | Chordata |
| Lớp (class) | Sauropsida |
| Bộ (ordo) | Squamata |
| Phân bộ (subordo) | SerpentesLinnaeus, 1758 |
| Cận bộ và họ | |
| |
Rắn là tên gọi chung để chỉ một nhóm các loài động vật bò sát ăn thịt, không có chân và thân hình tròn dài (hình trụ), thuộc phân bộ Serpentes, có thể phân biệt với các loài thằn lằn không chân bằng các đặc trưng như không có mí mắt và tai ngoài. Giống như các loài bò sát có vảy (Squamata) khác, rắn là động vật có xương sống, có màng ối, ngoại nhiệt với các lớp vảy xếp chồng lên nhau che phủ cơ thể. Nhiều loài rắn có sọ với nhiều khớp nối hơn các tổ tiên là động vật dạng thằn lằn của chúng, cho phép chúng nuốt các con mồi to lớn hơn nhiều so với đầu chúng với các quai hàm linh động cao. Để phù hợp với cơ thể thon và hẹp của mình, các cơ quan có cặp đôi của rắn được bố trí theo kiểu cái này nằm phía trước cái kia thay vì ngang hàng ở hai bên, và phần lớn các loài rắn chỉ có 1 phổi hoạt động. Một vài loài vẫn duy trì một đai chậu với 1 cặp vuốt dạng vết tích ở một trong hai bên của lỗ huyệt.
Các loài rắn còn sinh tồn đã được tìm thấy trên gần như mọi châu lục (ngoại trừ châu Nam Cực), trong lòng các đại dương như Ấn Độ Dương và Thái Bình Dương, và trên phần lớn các khối lục địa nhỏ hơn — các ngoại lệ bao gồm một số đảo lớn như Ireland và New Zealand, và nhiều đảo nhỏ trong Đại Tây Dương và Trung Thái Bình Dương.[3] Trên 20 họ rắn hiện nay đang được công nhận, bao gồm khoảng 500 chi với khoảng 3.400 - 3.550 loài.[2][4] Kích thước của chúng biến động từ nhỏ, như rắn chỉ (Leptotyphlops carlae) chỉ dài khoảng 10 cm cho tới lớn như trăn gấm (Python reticulatus) dài tới 8,7 m.[5][6] Loài tìm thấy ở dạng hóa thạch là Titanoboa cerrejonensis dài tới 15 m. Người ta cho rằng rắn đã tiến hóa từ các loài động vật dạng thằn lằn hoặc là sống đào bới hoặc là thủy sinh trong giai đoạn Creta giữa, và các hóa thạch sớm nhất đã biết có niên đại khoảng 112 Ma. Sự đa dạng của rắn đã xuất hiện trong thế Paleocen (khoảng 66 tới 56 Ma).
Phần lớn các loài rắn không có nọc độc, còn những loài nào có nọc độc thì chủ yếu sử dụng nó vào việc giết chết hay khuất phục con mồi thay vì để phòng vệ. Một số loài có nọc độc mạnh tới mức đủ gây ra vết thương đau nhức hay gây tử vong cho con người. Các loài rắn không nọc độc hoặc là nuốt sống con mồi hoặc là giết nó bằng cách quấn và vặn xiết.
Từ nguyên
[sửa | sửa mã nguồn]Trong tiếng Việt, từ rắn bắt nguồn từ *p-səɲʔ trong tiếng Việt-Mường nguyên thủy, chính nó lại bắt nguồn từ *(b)saɲʔ trong tiếng Nam Á nguyên thủy.
Trong tiếng Anh, từ snake bắt nguồn từ snaca trong tiếng Anh thượng cổ, chính nó phái sinh từ *snak-an- trong tiếng German nguyên thủy, gốc từ tiếng Proto-Ấn-Âu là *(s)nēg-o- 'bò, trườn', cũng chính là gốc của từ sneak 'lén lút' và từ nāgá 'rắn' trong tiếng Phạn.[7] Một từ nữa cho rắn trong tiếng Anh là serpent được mượn từ tiếng Pháp, bắt nguồn từ tiếng Ấn-Âu nguyên thủy là *serp- 'trườn',[8] cũng chính là gốc của từ ἕρπω (hérpō) 'Tôi bò' trong tiếng Hy Lạp cổ đại.
Tiến hóa
[sửa | sửa mã nguồn]Tổng quan phát sinh chủng loài các nhóm còn sinh tồn
Lưu ý: Biểu đồ này chỉ chỉ ra các mối quan hệ chứ không chỉ ra khoảng thời gian tạo nhánh tiến hóa..[9][10] |
Hồ sơ hóa thạch rắn là khá nghèo nàn do các bộ xương rắn thông thường là nhỏ và dễ gãy, làm cho sự hóa thạch là không phổ biến. Các hóa thạch có thể được nhận dạng như là rắn (mặc dù thường vẫn duy trì các chi sau) lần đầu tiên xuất hiện trong hồ sơ hóa thạch trong kỷ Creta.[11] Các hóa thạch rắn sớm nhất đã biết được tìm thấy tại các di chỉ trong khu vực Utah và Algérie, đại diện tương ứng cho các chi Coniophis và Lapparentophis. Các hóa thạch này được cho là có niên đại tương ứng với tầng Alba hay tầng Cenomanum (dù không chắc chắn lắm) vào cuối kỷ Creta, khoảng 112 tới 94 Ma. Tuy nhiên, niên đại thậm chí còn sớm hơn đã từng được đề xuất cho một trong các di chỉ tại Algérie, có thể cổ tới mức tương ứng với tầng Apt, 125 tới 112 Ma.[12]
Dựa theo giải phẫu học so sánh, người ta có sự đồng thuận cho rằng rắn là hậu duệ của một nhóm thằn lằn.[13]:11[14] Người ta cũng cho rằng các loài trăn trong các họ Pythonidae và Boidae là các nhóm nguyên thủy nhất trong số các loài rắn hiện đại—chúng có các chi sau dạng vết tích: các ngón chân nhỏ xíu có vuốt gọi là các cựa hậu môn, được chúng sử dụng để bấu víu trong khi giao phối.[6]:11[15] Các họ Leptotyphlopidae và Typhlopidae cũng có các dấu tích của đai chậu, đôi khi xuất hiện như là các chỗ lồi ra dạng chất sừng khi nhìn thấy.
Tất cả các loài rắn hiện đã biết đều không có chi trước. Điều này là do sự tiến triển của các gen Hox, kiểm soát sự tạo hình chi. Cột sống của tổ tiên chung của các loài rắn, giống như phần lớn các loài động vật bốn chân khác, có các chuyên biệt hóa khu vực bao gồm các đốt sống cổ, ngực, thắt lưng, cùng (chậu) và đuôi. Ngay từ ban đầu trong tiến hóa của rắn thì sự biểu lộ các gen Hox trong phần cột sống chịu trách nhiệm cho sự phát triển đốt sống ngực đã chiếm ưu thế. Kết quả là tất cả các đốt sống phía trước các chồi chi sau (khi có mặt) đều có cùng một nhận dạng giống như kiểu đốt sống ngực (ngoại trừ từ đốt sống đội, đốt sống trục và các đốt sống cổ 1–3). Nói cách khác, phần lớn bộ xương của rắn có thể coi là một cái ngực kéo cực kỳ dài. Các xương sườn chỉ được tìm thấy trên các đốt sống ngực. Các đốt sống cổ, thắt lưng và khung chậu bị giảm đi rất nhiều về lượng (chỉ có 2–10 đốt sống thắt lưng và chậu), trong khi chỉ có các dấu tích ngắn của các đốt sống đuôi. Tuy nhiên, đuôi vẫn đủ dài để có công dụng quan trọng ở nhiều loài, và nó bị biến đổi ở một số loài sống dưới nước hay leo trèo trên cây.
Các loài rắn hiện đại đã đa dạng hóa mạnh trong thế Paleocen. Điều này diễn ra cùng với phân tỏa thích nghi của động vật có vú, ngay sau sự kiện tuyệt chủng của khủng long phi chim. Các loài rắn thuộc họ Colubridae, một trong các nhóm rắn phổ biến nhất, dã trở nên cực kỳ đa dạng do thức ăn của chúng là động vật gặm nhấm, một nhóm động vật có vú đặc biệt thành công.
Nguồn gốc
[sửa | sửa mã nguồn]Nguồn gốc của rắn vẫn là một vấn đề chưa được giải quyết. Có hai giả thuyết chính cạnh tranh lẫn nhau về nguồn gốc của rắn.
Giả thuyết thằn lằn đào bớiCó chứng cứ hóa thạch cho thấy rắn có thể đã tiến hóa từ thằn lằn sống kiểu đào bới, chẳng hạn như Varanidae (hoặc một nhóm tương tự) trong kỷ Creta.[16] Một loài rắn hóa thạch cổ, Najash rionegrina, là một động vật đào bới có hai chân với xương cùng, và sống hoàn toàn trên cạn.[17] Một loài còn sinh tồn tương tự như tổ tiên giả định này là kỳ đà không tai (Lanthanotus borneensis) ở Borneo (mặc dù nó là động vật bán thủy sinh).[18] Các loài sống ngầm dưới lòng đất đã tiến hóa để có một cơ thể thuôn dạng khí động học phục vụ cho việc đào bới, và cuối cùng đã mất đi các chi.[18] Theo giả thuyết này, các đặc trưng như độ trong suốt, các mí mắt hợp nhất (vảy mắt) và sự mất đi các tai ngoài đã tiến hóa để đương đầu với các khó khăn trong cuộc sống đào bới, như các giác mạc bị xây sát và chất bẩn lọt vào tai.[16][18] Một số loài rắn nguyên thủy còn có các chi sau, nhưng các xương chậu của chúng không có kết nối trực tiếp tới cột sống. Chúng bao gồm các loài hóa thạch như Haasiophis, Pachyrhachis và Eupodophis, đều có niên đại hơi cổ hơn Najash.[15]

Một giả thuyết khác, dựa vào hình thái học, cho rằng tổ tiên của rắn là có họ hàng với thương long (Mosasauridae)—một nhóm bò sát thủy sinh đã tuyệt chủng, từng sinh sống trong kỷ Creta—tới lượt chúng, thương long lại được coi là có nguồn gốc từ thằn lằn dạng kỳ đà (Varanoidea).[14] Theo giả thuyết này thì các mí mắt trong suốt và hợp nhất của rắn được cho là đã tiến hóa để chống lại các điều kiện biển cả (dịch giác mạc bị mất qua thẩm thấu), và các tai ngoài bị mất do nó là vô dụng trong môi trường nước. Điều này cuối cùng dẫn tới con vật tương tự như rắn biển ngày nay. Vào cuối kỷ Creta, rắn đã tái chiếm đất liền và tiếp tục đa dạng hóa thành rắn ngày nay. Các dấu tích rắn hóa thạch được biết đến trong các trầm tích biển thuộc thời kỳ đầu của giai đoạn Creta muộn là phù hợp với giả thuyết này; đặc biệt là chúng đều cổ hơn so với loài Najash rionegrina sống trên cạn. Cấu trúc hộp sọ tương tự, các chi tiêu giảm hay không có, và các đặc trưng giải phẫu khác được tìm thấy ở cả thương long lẫn rắn dẫn tới mối tương quan miêu tả theo nhánh rõ ràng, mặc dù một vài đặc trưng này cũng được chia sẻ với thằn lằn dạng kỳ đà.
Các nghiên cứu di truyền gần đây đã chỉ ra rằng rắn không có quan hệ họ hàng gần với kỳ đà như người ta đã từng một thời tin tưởng—và vì thế cũng không gần với thương long, tổ tiên được đề xuất trong kịch bản thủy sinh trong quá trình tiến hóa của chúng. Tuy nhiên, có nhiều chứng cứ liên kết thương long với rắn hơn là với kỳ đà. Các dấu tích phân mảnh tìm thấy trong thời kỳ từ kỷ Jura và Creta sớm chỉ ra các hồ sơ hóa thạch sâu hơn cho các nhóm này, và điều này có thể sẽ bác bỏ cả hai giả thuyết vừa đề cập.
Phân bố
[sửa | sửa mã nguồn]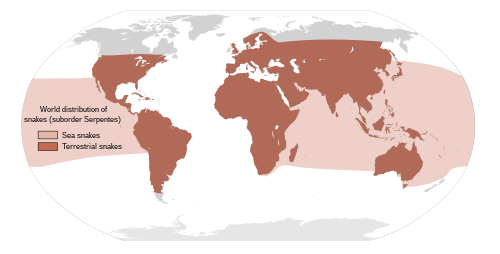
Có khoảng 3.500 loài rắn phân bố từ vùng phía bắc tới vòng Bắc Cực tại Scandinavia cho tới phía nam tại Australia.[14] Rắn có thể được thấy trên gần như mọi châu lục (trừ châu Nam Cực), trong lòng đại dương, cũng như cao tới cao độ 4.900 m (16.000 ft) trong khu vực dãy núi Himalaya ở châu Á.[14][19]:143 Có nhiều hòn đảo không có rắn, như Ireland, Iceland và New Zealand[3][19] (mặc dù vùng biển ven New Zealand thỉnh thoảng vẫn thấy có rắn đẻn sọc dưa (Pelamis platura) và rắn cạp nia biển (Laticauda colubrina)[20]
Phân loại
[sửa | sửa mã nguồn]Tất cả các loài rắn hiện đại được gộp nhóm trong phân bộ Serpentes trong phân loại học Linnaeus, một phần của bộ Squamata (bò sát có vảy), mặc dù vị trí chính xác của chúng trong phạm vi bò sát có vảy vẫn là điều gây mâu thuẫn.[2]
Người ta thường chia phân bộ này thành 2 cận bộ là Alethinophidia và Scolecophidia.[2] Sự chia tách này dựa theo các đặc trưng hình thái học và sự tương đồng trình tự DNA ti thể. Alethinophidia đôi khi được chia tách tiếp thành Henophidia và Caenophidia, với nhóm thứ hai bao gồm các loài rắn dạng "rắn nước" (Colubridae, Viperidae, Elapidae, Hydrophiidae và Attractaspididae) và Acrochordidae, trong khi các họ rắn dạng Alethinophidia còn lại tạo thành Henophidia.[21] Trong khi không còn loài nào sinh tồn ngày nay nhưng Madtsoiidae, một họ bao gồm các loài rắn khổng lồ; nguyên thủy; giống như trăn, đã từng sinh sống cho tới khoảng 50.000 năm trước tại Australia, với các chi như Wonambi.
Có nhiều tranh luận liên quan tới hệ thống học trong phạm vi nhóm này. Chẳng hạn, nhiều nguồn coi Boidae và Pythonidae chỉ là một họ trong khi quan hệ họ hàng của chúng không thực sự gần, trong khi lại coi Elapidae (rắn hổ) và Hydrophiidae (rắn biển) là các họ tách biệt vì các lý do thực tiễn trong khi quan hệ của chúng là rất gần gũi.
Các nghiên cứu phân tử gần đây hỗ trợ cho tính đơn ngành của nhánh chứa các loài rắn hiện đại, cũng như của hai nhánh Scolecophidia (Anomalepididae + (Leptotyphlopidae + (Gerrhopilidae + (Xenotyphlopidae + Typhlopidae)))) và Alethinophidia (Anilius + Alethinophidia phần lõi (Uropeltidae (Cylindrophis + Anomochilus + Uropeltinae) + Macrostomata (Tropidophiinae + Pythonidae + Caenophidia + Booidea (Boidae + Ungaliophiinae))).[9]
Các họ
[sửa | sửa mã nguồn]| Cận bộ Alethinophidia 15 họ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Họ[2] | Tác giả[2] | Số chi[2] | Số loài[2] | Tên thường gọi | Phân bố[22] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acrochordidae | Bonaparte, 1831 | 1 | 3 | Rắn rằm ri cóc | Tây Ấn Độ và Sri Lanka qua vùng nhiệt đới Đông Nam Á tới Philippines, về phía nam qua nhóm đảo Indonesia/Malaysia tới Timor, về phía đông qua New Guinea tới vùng duyên hải phía bắc Australia tới đảo Mussau, quần đảo Bismark và đảo Guadalcanal trong quần đảo Solomon. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Aniliidae | Stejneger, 1907 | 1 | 1 | Rắn san hô giả | Nhiệt đới Nam Mỹ. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Anomochilidae | Cundall, Wallach, 1993 | 1 | 2 | Rắn ống lùn | Tây Malaysia và trên đảo Sumatra của Indonesia. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atractaspididae | Günther, 1858 | 12 | 64 | Burrowing asps | châu Phi và Trung Đông.[6][23][24] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Boidae | Gray, 1825 | 8 | 43 | Trăn | Bắc, Trung và Nam Mỹ, Caribe, đông nam châu Âu và Tiểu Á, Bắc, Trung và Đông Phi, Madagascar và đảo Reunion, bán đảo Ả Rập, Trung và Tây Nam Á, Ấn Độ và Sri Lanka, Moluccas và New Guinea cho tới Melanesia và Samoa. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Bolyeriidae | Hoffstetter, 1946 | 2 | 2 | Trăn đảo Round, rắn hàm chẻ | Mauritius. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Colubridae | Oppel, 1811 | 304[4] | 1938[4] | Rắn điển hình | Phổ biến rộng khắp trên mọi châu lục, trừ châu Nam Cực[25] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cylindrophiidae | Fitzinger, 1843 | 1 | 8 | Rắn trun, rắn hai đầu | Sri Lanka về phía đông qua Myanma, Thái Lan, Campuchia, Việt Nam và bán đảo Mã Lai xa về phía đông tới quần đảo Aru ngoài khơi tây nam New Guinea. Cũng có ở miền nam Trung Quốc (Phúc Kiến, Hồng Kông và Hải Nam) và Lào. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Elapidae | Boie, 1827 | 61 | 235 | Rắn hổ | Trên cạn, khắp thế giới trong khu vực nhiệt đới và cận nhiệt đới, trừ châu Âu. Nếu gộp cả rắn biển thì có trong Ấn Độ Dương và Thái Bình Dương.[26] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Loxocemidae | Cope, 1861 | 1 | 1 | Rắn đào bới Mexico | Dọc theo các sườn núi ven Thái Bình Dương từ Mexico về phía nam tới Costa Rica. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Pythonidae | Fitzinger, 1826 | 8 | 26 | Trăn | châu Phi hạ Sahara, Ấn Độ, Myanma, Hoa Nam, Đông Nam Á và từ Philippines về phía đông nam qua Indonesia tới New Guinea và Australia. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tropidophiidae | Brongersma, 1951 | 4 | 22 | Trăn lùn | Từ nam Mexico và Trung Mỹ về phía nam tới tây bắc Nam Mỹ (lưu vực sông Amazon) ở Colombia, Ecuador và Peru, cũng như ở tây bắc và đông nam Brasil. Cũng có ở Tây Ấn. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Uropeltidae | Müller, 1832 | 8 | 47 | Rắn đuôi khiên | Nam Ấn Độ và Sri Lanka. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Viperidae | Oppel, 1811 | 32 | 224 | Rắn lục | châu Mỹ, châu Phi và đại lục Á-Âu. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Xenopeltidae | Bonaparte, 1845 | 1 | 2 | Rắn mống | Đông Nam Á từ quần đảo Andaman và Nicobar về phía đông qua Myanma tới Hoa Nam, Thái Lan, Lào, Campuchia, Việt Nam, bán đảo Mã Lai và Đông Ấn tới Sulawesi, cũng như tại Philippines. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cận bộ Scolecophidia 5 họ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Họ[2] | Tác giả[2] | Số chi[2] | Số loài[2] | Tên thường gọi | Phân bố[22] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anomalepidae | Taylor, 1939 | 4 | 17 | Rắn mù nguyên thủy | Từ miền nam Trung Mỹ tới tây bắc Nam Mỹ. Các quần thể rời rạc tại đông bắc và đông nam Nam Mỹ. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Leptotyphlopidae | Stejneger, 1892 | 2 | 120 | Rắn mù thân mảnh | châu Phi, Tây Á từ Thổ Nhĩ Kỳ tới tây bắc Ấn Độ, trên đảo Socotra, từ tây nam Hoa Kỳ về phía nam qua Mexico và Trung Mỹ tới Nam Mỹ, nhưng không có trong khu vực miền núi cao trong dãy Andes. Tại Nam Mỹ ven Thái Bình Dương chúng có mặt xa về phía nam tới vùng duyên hải nam Peru, còn ở phía Đại Tây Dương thì xa tới Uruguay và Argentina. Tại khu vực Caribe chúng có mặt tại Bahamas, Hispaniola và Tiểu Antilles. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Typhlopidae | Merrem, 1820 | 4 | 240 | Rắn mù điển hình, rắn giun | Rộng khắp thế giới trong khu vực nhiệt đới và cận nhiệt đới, cụ thể tại châu Phi, Madagascar, châu Á, Australia, các đảo trên Thái Bình Dương, nhiệt đới châu Mỹ và tại đông nam châu Âu. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gerrhopilidae | Vidal, Wynn, Donnellan & Hedges, 2010 | 1 | 15 | Rắn mù, rắn giun | Nam Á, Đông Nam Á, New Guinea. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Xenotyphlopidae | Vidal, Wynn, Donnellan & Hedges, 2010 | 1 | 2 | Rắn mù Madagascar | Madagascar. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sinh học
[sửa | sửa mã nguồn]
Bộ xương
[sửa | sửa mã nguồn]Bộ xương của phần lớn các loài rắn chỉ bao gồm sọ, xương móng, cột sống và các xương sườn, mặc dù các loài rắn thuộc nhánh Henophidia vẫn còn các vết tích của khung chậu và các chi sau. Sọ của rắn bao gồm một hộp sọ đặc và hoàn hảo, và nhiều xương khác chỉ gắn vào nó một cách lỏng lẻo, cụ thể là các xương hàm có độ linh động cao, tạo thuận lợi cho việc bắt giữ và nuốt các con mồi to lớn. Các bên trái và phải của hàm dưới chỉ được nối bằng một dây chằng dễ uốn vào các chóp trước, cho phép chúng tách rộng ra, trong khi các chóp sau của các xương hàm dưới nối khớp với xương vuông, tạo thêm tính cơ động. Các xương của hàm dưới và xương vuông cũng có thể thu nhận các rung động phát sinh từ mặt đất.[27] Do các bên của xương hàm có thể di chuyển độc lập với nhau nên khi rắn đặt các quai hàm của nó trên một bề mặt thì nó có thính giác lập thể nhạy cảm giúp nó có thể phát hiện vị trí của con mồi. Lộ trình quai hàm-xương vuông-xương bàn đạp có khả năng phát hiện các rung động ở cấp độ angstrom, mặc cho sự thiếu vắng tai ngoài và cơ chế xương nhỏ thính giác của khớp trở kháng được sử dụng ở các động vật có xương sống khác để thu nhận các rung động từ không trung.[28][29] Xương móng là một xương nhỏ nằm ở phía sau và trên mặt bụng của sọ, trong khu vực 'cổ', có tác dụng như một bộ phận gắn kết cho các cơ của lưỡi rắn, cũng giống như ở tất cả các động vật bốn chân khác.
Cột sống của rắn bao gồm khoảng 200 tới 400 (hoặc hơn) đốt sống. Các đốt sống đuôi là tương đối ít về số lượng (thường không quá 20% tổng số đốt sống) và không có xương sườn, trong khi các đốt sống phần thân đều có 2 xương sườn nối khớp với chúng. Các đốt sống đều có các phần lồi ra cho phép có sự kết nối với các cơ khỏe giúp cho việc di chuyển không cần tới chân. Sự tự đứt đuôi, một đặc trưng ở một số loài thằn lằn nói chung lại không có ở phần lớn các loài rắn.[30] Sự tự đứt đuôi ở rắn là hiếm thấy và là liên đốt sống, không giống như ở thằn lằn là cơ chế nội đốt sống—nghĩa là sự ngắt đuôi xảy ra dọc theo một mặt phẳng đứt gãy được xác định trước và có sẵn trên đốt sống.[31][32]
Ở một số loài rắn, đáng chú ý là các họ Boidae và Pythonidae, có các vết tích của các chi sau dưới dạng một cặp cựa khung chậu. Các phần thò ra nhỏ và giống như vuốt này ở mỗi bên của lỗ huyệt là phần bên ngoài của phần xương chi sau dạng vết tích, bao gồm các phần sót lại của xương chậu và xương đùi.
Nội tạng
[sửa | sửa mã nguồn]Tim rắn được bao bọc trong một túi, gọi là màng ngoài tim, nằm ở chỗ rẽ đôi của phế quản. Tuy nhiên tim rắn có thể di chuyển vòng quanh do không có cơ hoành. Sự điều chỉnh này bảo vệ tim khỏi bị tổn thương khi con mồi nuốt vào là to lớn và trượt dọc theo thực quản. Lá lách gắn với túi mật và tụy, giúp lọc máu. Tuyến ức nằm trong mô mỡ phía trên tim, chịu trách nhiệm sản sinh các tế bào miễn dịch trong máu. Hệ tim mạch của rắn cũng là độc đáo ở chỗ có một hệ thống cổng thận, trong đó máu từ đuôi rắn di chuyển qua thận trước khi trở về tim.[33]
Phổi trái dạng vết tích thường là nhỏ hoặc không có, do cơ thể hình ống của rắn đòi hỏi mọi cơ quan phải dài và mỏng.[33] Ở phần lớn các loài rắn chỉ phổi phải là hoạt động. Phổi phải này bao gồm hai phần: phần trước có mạch máu dẫn tới, còn phần sau không có tác dụng trong trao đổi khí.[33] 'Phổi dạng túi' này được sử dụng cho các mục đích thủy tĩnh để điều chỉnh sức nổi ở một số loài rắn thủy sinh và chức năng của nó đối với các loài rắn sống trên cạn thì vẫn chưa rõ.[33] Nhiều cơ quan có cặp đôi, như thận hay cơ quan sinh dục, được sắp xếp xen kẽ trong cơ thể, với một bộ phận ('trái' hay 'phải') của cơ quan này nằm ngay phía trước bộ phận còn lại ('phải' hay 'trái') của chính cơ quan đó.[33] Rắn không có hạch bạch huyết.[33]
Kích thước
[sửa | sửa mã nguồn]
Loài trăn đã tuyệt chủng Titanoboa cerrejonensis dài tới 12–15 m (39–49 ft). Trong khi đó, trong số loài rắn còn sinh tồn thì loài được coi là dài nhất là trăn gấm (Python reticulatus) với chiều dài lên tới khoảng 9 m (30 ft), còn loài được coi là nặng nhất là trăn anaconda xanh (Eunectes murinus) dài khoảng 7,5 m (25 ft)[34] và nặng tới 97,5 kg.[35]
Ngược lại, ở đầu kia của dải kích thước, loài rắn nhỏ nhất còn sinh tồn là rắn giun Barbados (Leptotyphlops carlae), với chiều dài chỉ khoảng 10 cm (4 inch).[36] Phần lớn các loài rắn là động vật tương đối nhỏ, dài khoảng 1 m (3 ft).[37]
Da
[sửa | sửa mã nguồn]
Da rắn được che phủ trong một lớp vảy sừng. Trái với ý niệm phổ biến cho rằng da rắn nhớt, có lẽ là do sự nhầm lẫn rắn với giun, trên thực tế da rắn nhẵn nhụi và khô. Phần lớn các loài rắn sử dụng các vảy bụng chuyên biệt hóa để di chuyển, bám lấy các bề mặt. Các vảy trên cơ thể rắn có thể là nhẵn nhụi, có gờ hay dạng hạt. Các mí mắt của rắn là các vảy sừng trong suốt, giống như "đeo kính", và các vảy này luôn luôn đóng kín, vì thế mà có thành ngữ "Thao láo như mắt rắn ráo".
Sự lột bỏ lớp vảy ở rắn gọi là lột xác. Trong trường hợp lột da ở rắn thì nguyên lớp da ngoài cùng bị lột bỏ.[38] Các vảy của rắn không phải là rời rạc mà là sự mở rộng của lớp biểu bì—vì thế chúng không bị lột tách biệt mà như là một lớp ngoài cùng hoàn hảo trong mỗi lần lột xác, giống như việc người ta lột mặt trong của cái tất ra bên ngoài.[39]
Hình dáng và số lượng vảy trên đầu, lưng và bụng thường là đặc trưng và được sử dụng vào mục đích phân loại. Các vảy được đặt tên chủ yếu theo vị trí của chúng trên cơ thể. Trong nhóm rắn "bậc cao" (Caenophidia), các vảy bụng rộng bản và các hàng vảy lưng tương ứng với các đốt sống, cho phép các nhà khoa học có thể đếm số lượng đốt sống mà không cần phải mổ xẻ.
Mắt rắn được che phủ bằng vảy trong suốt chứ không phải mí mắt có thể chuyển động. Vì thế mắt rắn luôn luôn mở, còn để ngủ thì võng mạc có thể khép lại hoặc rắn giấu đầu vào phần thân đã cuộn tròn.
Lột xác
[sửa | sửa mã nguồn]
Sự lột xác (hay lột da) ở rắn phục vụ cho một loạt các chức năng. Trước hết,lớp da ngoài cũ kỹ và đã bị mòn được thay thế; thứ hai, nó giúp loại bỏ các động vật ký sinh như ve hay bét. Việc làm mới lớp da nhờ lột xác được cho là giúp đỡ cho quá trình phát triển ở một số động vật như côn trùng; tuy nhiên điều này có đúng như vậy hay không ở rắn thì vẫn là điều còn gây tranh cãi[39][40]
Sự lột da xảy ra theo chu kỳ trong suốt cuộc đời rắn. Trước khi lột, rắn ngừng ăn uống và thường di chuyển tới hay ẩn nấp tại nơi an toàn. Ngay trước khi lột, lớp da ngoài trở nên xỉn màu và khô và hai mắt thì mờ đục hay xám xỉn. Mặt trong của lớp da cũ hóa lỏng làm cho lớp da cũ tách ra khỏi lớp da mới nằm ngay dưới nó. Sau vài ngày thì mắt trong trở lại và con rắn "trườn" ra khỏi lớp da cũ của nó. Lớp da cũ đứt ở phần gần miệng và rắn lách ra ngoài, hỗ trợ quá trình lột bỏ bằng cách cọ xát vào các bề mặt thô nhám. Trong nhiều trường hợp thì lớp da cũ bị lột ngược dọc theo thân từ đàu tới đuôi thành một tấm, giống như sự lột tất ngược từ trong ra ngoài. Lớp da mới, lớn hơn và sáng màu hơn đã được hình thành ngay phía dưới.[39][41]
Những con rắn già chỉ lột da 1 tới 2 lần mỗi năm, nhưng những con rắn non còn đang lớn thì có thể lột da tới 4 lần mỗi năm. Lớp da bị loại bỏ giữ nguyên vẹn dấu vết của kiểu vảy, và thường thì có thể dùng nó để nhận dạng loài rắn đó nếu như lớp da bị loại bỏ này còn giữ được khá nguyên vẹn.[39] Sự lột da theo chu kỳ này là lý do chính để biến rắn thành một biểu tượng của điều trị bệnh tật và y học, như trong biểu tượng gậy Asclepius (xà trượng)[42]
Giác quan
[sửa | sửa mã nguồn]
Nọc
[sửa | sửa mã nguồn] Xem thêm: Nọc rắn
Rắn hổ, rắn lục và các loài họ hàng gần của chúng sử dụng nọc để làm tê liệt hay giết chết con mồi. Nọc của chúng thực tế là nước bọt đã biến đổi, được tiết ra theo các răng nọc.[6]:243 Các răng nọc của các loài rắn có nọc độc 'bậc cao' như rắn lục hay rắn hổ là các răng rỗng để tiêm nọc hiệu quả hơn, trong khi răng nọc của các loài rắn với răng nọc ở phía sau như rắn cây châu Phi (Dispholidus typus) đơn giản chỉ là một khía rãnh trên rìa sau để nọc chảy vào vết thương. Nọc rắn là đặc trưng chủ yếu dành cho săn mồi—vai trò của nó trong phòng vệ chỉ là thứ cấp.[6]:243
Nọc, cũng giống như các loại dịch tiết dạng nước bọt khác, là chất tiền tiêu hóa để bắt đầu sự phân tách thức ăn thành các hợp chất hòa tan, hỗ trợ cho sự tiêu hóa. Ngay cả những cú cắn của rắn không có nọc độc (giống như của cú cắn của bất kỳ động vật nào) đều gây ra tổn thương mô.[6]:209
Một số loài chim, thú hay rắn (chẳng hạn như rắn vua (Lampropeltis)) nhất định với con mồi là những con rắn có nọc đều phát triển khả năng đề kháng hay thậm chí là miễn nhiễm với một số loại nọc độc nhất định.[6]:243 Các loài rắn có nọc nằm trong 3-4 họ rắn, nhưng chúng không và không thể hợp lại thành một nhóm phân loại chính thức nào để dùng trong phân loại học.
Thuật ngữ rắn độc đa phần là thiếu chính xác. Chất độc (hay độc tố) là những chất được hít hay nuốt vào, trong khi nọc được tiêm vào.[45] Tuy nhiên ở đây vẫn có 2 ngoại lệ: Rắn hoa cỏ (Rhabdophis) cô lập các chất độc từ những con cóc mà nó ăn, sau đó tiết các chất độc này từ các tuyến gáy để ngăn cản những kẻ săn mồi, và một quần thể nhỏ rắn garter (Thamnophis) ở Oregon duy trì đủ lượng độc tố trong gan của chúng từ cá cóc/sa giông (Pleurodelinae) mà chúng ăn vào để tự bản thân trở thành đủ độc đối với những kẻ săn mồi nhỏ bé tại khu vực này (như quạ hay cáo).[46]
Nọc rắn là các hỗn hợp phức tạp của các protein, và được lưu trữ trong các tuyến độc ở phía sau đầu.[46] Ở tất cả các loài rắn có nọc, các tuyến này đổ thông qua các ống dẫn vào các răng rỗng hay răng có khía ở hàm trên.[6]:243[45] Các protein này có thể là hỗn hợp thuộc các nhóm neurotoxin (tác động thần kinh), hemotoxin (tác động hệ tuần hoàn), cytotoxin (tác động tế bào), bungarotoxin (độc tố cạp nia) và nhiều loại độc tố khác có tác động tới cơ thể theo cách thức khác nhau.[45] Gần như tất cả các loại nọc rắn đều chứa hyaluronidaza, một enzym đảm bảo sự khuếch tán nhanh của nọc.[6]:243
Các loại rắn có nọc sử dụng độc tố hoại máu (hemotoxin) thường có các răng nọc ở phía trước miệng chúng, giúp chúng dễ dàng hơn trong việc tiêm nọc vào nạn nhân của chúng.[45] Một số loài rắn sử dụng neurotoxin, như rắn rào cây (Boiga dendrophila) có các răng nọc ở phía sau miệng chúng với các răng này uốn cong ngược về phía sau.[47] Điều này gây khó khăn cho cả con rắn trong việc sử dụng nọc của nó lẫn cho các nhà khoa học trong việc lấy nọc rắn.[45] Tuy nhiên, các loài rắn hổ, như rắn hổ mang hay cạp nia lại là xẻ khía trước— ở hàm trên của chúng có răng nọc rỗng trỏ xuống phía dưới nhưng không thể dựng đứng lên để hướng về phía trước miệng của chúng, và không thể "đâm" như rắn lục/rắn vipe. Chúng phải thực sự cắn nạn nhân.[6]:242
Gần đây người ta đề xuất cho rằng tất cả các loài rắn đều có thể là rắn có nọc ở một mức độ nào đó, với các loài rắn vô hại có nọc yếu và không có răng nọc.[48] Phần lớn các loài rắn hiện tại được gán nhãn "không độc" có thể vẫn được coi là vô hại theo thuyết này, do chúng hoặc là không có phương thức truyền nọc hoặc là không đủ khả năng truyền một lượng nọc đủ để gây nguy hiểm cho con người. Thuyết này đưa ra định đề rằng rắn có thể đã tiến hóa từ một tổ tiên chung là động vật dạng thằn lằn có nọc—và rằng các loài thằn lằn có nọc như thằn lằn độc Gila (Heloderma suspectum), thằn lằn độc Mexico (Heloderma horridum), kỳ đà (Varanidae), cũng như thương long đã tuyệt chủng cũng có thể đã chuyển hóa. Chúng chia sẻ cùng một nhánh có nọc này với nhiều loài bò sát hai cung khác trong nhánh Sauria.
Các loài rắn có nọc độc về cơ bản được phân loại trong 2 họ:
- Elapidae – Họ Rắn hổ, bao gồm rắn hổ mang (Naja), rắn hổ mang chúa (Ophiophagus hannah), rắn cạp nong (Bungarus fasciatus), rắn cạp nia (Bungarus spp.), rắn mamba (Dendroaspis spp.), rắn đầu đồng Úc (Austrelaps), rắn biển (Hydrophiinae) và rắn san hô (các chi Leptomicrurus, Micruroides, Micrurus), rắn lá khô (Calliophis).[47]
- Viperidae – Họ Rắn lục, bao gồm rắn lục (Trimeresurus spp.), rắn lục đầu bạc (Azemiops feae), rắn vipe (Viperinae), rắn đuôi chuông (các chi Crotalus, Sistrurus), rắn đầu đồng Mỹ (Agkistrodon contortrix), rắn miệng bông (Agkistrodon piscivorus) và rắn chúa bụi (Lachesis spp.).[47]
Họ thứ ba chứa một số loài rắn răng nọc sau:
- Colubridae – Họ Rắn nước, bao gồm một số loài như rắn cây châu Phi (Dispholidus typus), rắn roi (Ahaetulla spp.), rắn rào (Boiga spp.), mặc dù không phải mọi loài trong họ Colubridae đều là rắn độc.[6]:209[47]
Tập tính
[sửa | sửa mã nguồn]Kiếm ăn và thức ăn
[sửa | sửa mã nguồn]


Tất cả các loài rắn đều là động vật chỉ toàn ăn thịt, với thức ăn của chúng là các động vật nhỏ như thằn lằn, chim, thú nhỏ, cá, côn trùng, ốc, các loài rắn khác cũng như trứng của các loại con mồi này[6][1].[14][49] Do rắn không thể cắn hay xé thức ăn ra thành từng miếng nên chúng phải nuốt chửng nguyên cả con mồi. Kích thước cơ thể rắn có ảnh hưởng lớn tới tập tính ăn uống của nó. Các loài rắn nhỏ hơn thì ăn các con mồi nhỏ hơn. Chẳng hạn, những con trăn non có thể khởi đầu công việc ăn uống chỉ với những con thằn lằn hay chuột và dần dần tiến tới việc ăn những con hươu hay linh dương nhỏ cũng như trưởng thành.
Quai hàm của rắn là một cấu trúc phức tạp. Trái với niềm tin thông thường rằng rắn có thể bị trật khớp các quai hàm của nó, trên thực tế rắn có hàm dưới rất linh hoạt, hai nửa của nó không bị gắn cứng nhắc, với một loạt các khớp nối khác trong sọ rắn, cho phép rắn có thể há miệng đủ to để nuốt toàn bộ con mồi, ngay cả khi con mồi này có đường kính lớn hơn bề rộng cơ thể của rắn.[50] Chẳng hạn, rắn ăn trứng châu Phi (Dasypeltis) có các quai hàm rất linh hoạt, thích nghi để ăn những quả trứng to hơn cả đường kính đầu nó.[6]:81 Các loài rắn này không có răng, nhưng có các chỗ lồi ra trên rìa trong của cột sống, được chúng sử dụng để bóp vỡ vỏ trứng khi nó được nuốt vào.[6]:81
Trong khi phần lớn các loài rắn ăn đủ mọi loại con mồi động vật thì vẫn có những loài chỉ chuyên ăn một loại thức ăn nhất định. Rắn hổ mang chúa và rắn bandy-bandy (Vermicella annulata) ở Australia chỉ ăn thịt các loài rắn khác. Loài rắn chuông có thể ăn cả những con rắn do chúng đẻ ra nếu như những con này quá yếu không thể tồn tại được, những con rắn mẹ đã ăn khoảng 11% số trứng là do rắn chuông bị cạn kiệt năng lượng sau khi sinh nên buộc phải tồn tại bằng cách ăn thịt những con quá yếu và những quả trứng không có khả năng nở con. Pareas iwesakii và các loài rắn ăn ốc khác trong phân họ Pareatinae của họ Colubridae có nhiều răng ở mé phải miệng chúng hơn ở mé trái do vỏ ốc mà chúng ăn thường xoắn theo chiều kim đồng hồ.[6]:184[51]
Một số loài rắn có cú cắn chứa nọc độc được chúng dùng để giết chết con mồi trước khi ăn thịt nó.[49][52] Một số loài rắn khác lại giết con mồi bằng cách quấn và siết chặt,[49] trong khi có những loài chỉ đơn giản là nuốt sống toàn bộ con mồi.[6]:81[49]
Sau khi ăn, rắn chuyển sang trạng thái nghỉ trong khi quá trình tiêu hóa diễn ra.[53] Tiêu hóa là một hoạt động mãnh liệt, đặc biệt sau khi nuốt con mồi lớn. Ở những loài chỉ thỉnh thoảng mới ăn thì toàn bộ ruột co nhỏ lại giữa các bữa ăn để tiết kiệm năng lượng. Hệ tiêu hóa sau đó sẽ 'phình to' tới sức chứa tối đa trong vòng 48 giờ sau khi ăn con mồi. Là động vật ngoại nhiệt ("máu lạnh"), nhiệt độ môi trường xung quanh có vai trò lớn trong sự tiêu hóa của rắn. Nhiệt độ lý tưởng đối với rắn trong quá trình tiêu hóa là khoảng 30 °C (86 °F). Năng lượng trao đổi chất trong sự tiêu hóa của rắn nhiều tới mức như ở rắn đuôi chuông Mexico (Crotalus durissus) thì nhiệt độ bề mặt cơ thể tăng lên tới 1,2 °C (2,2 °F) trong quá trình tiêu hóa.[54] Vì điều này, một con rắn bị quấy nhiễu ngay sau khi ăn thường sẽ nôn con mồi ra để có thể thoát khỏi mối đe dọa mà nó đã nhận thấy được. Khi không bị quấy nhiễu thì quá trình tiêu hóa có hiệu quả cao nhất, với các enzym tiêu hóa của rắn hòa tan và hấp thụ mọi thứ, ngoại trừ lông và móng vuốt của con mồi bị bài tiết ra ngoài dưới dạng chất thải.
Vận động
[sửa | sửa mã nguồn]Không có chân không gây ra cản trở nào đối với sự vận động của rắn. Chúng đã phát triển một vài cách thức vận động khác nhau để giải quyết vấn đề trong từng môi trường cụ thể. Không giống như các dáng đi của các động vật có chân, là thứ tạo thành một thể liên tục, mỗi cách thức vận động của rắn là rời rạc và khác biệt với các cách thức còn lại; các chuyển tiếp giữa các cách thức là đột ngột.[55][56]
Chuyển động sóng ngang
[sửa | sửa mã nguồn] Bài chi tiết: Vận động ngoằn ngoèoChuyển động sóng ngang là cách thức duy nhất trong vận động trong môi trường nước, và cũng là cách thức vận động trên cạn phổ biến nhất.[56] Trong cách thức này, cơ thể rắn lần lượt uốn cong sang trái và sang phải, tạo ra một chuỗi các "sóng" chuyển động về phía sau.[55] Trong khi kiểu di chuyển này có vẻ là nhanh, nhưng hiếm khi thấy rắn di chuyển nhanh hơn 2 lần chiều dài cơ thể của nó mỗi giây, thường là nhỏ hơn thế.[57] Cách thức di chuyển này có cùng một tiêu hao năng lượng ròng (lượng calo tiêu hao trên mỗi mét di chuyển) như kiểu chạy của những con thằn lằn có cùng khối lượng.[58]
Trên cạn
[sửa | sửa mã nguồn]Chuyển động sóng ngang trên cạn là cách thức vận động trên cạn phổ biến nhất đối với phần lớn các loài rắn.[55] Trong cách thức này, các sóng chuyển động về phía sau đẩy vào các tiếp điểm trong môi trường, như các hòn đá, các cành cây hay các chỗ mấp mô trên mặt đất v.v.[55] Mỗi vật thể trong môi trường này tới lượt mình lại sinh ra một phản lực hướng về phía trước và về phía đường trung bình của con rắn, tạo ra lực đẩy về phía trước trong khi các thành phần ngang bị triệt tiêu.[59] Vận tốc của di chuyển này phụ thuộc vào mật độ các điểm đẩy trong môi trường, trong khi mật độ trung bình khoảng 8 dọc theo chiều dài thân rắn là lý tưởng.[57] Vận tốc truyền sóng chính xác bằng vận tốc của con rắn, và kết quả là mọi điểm trên thân rắn đi theo đường của điểm ngay trước nó, cho phép con rắn di chuyển qua thảm thực vật rất rậm rạp hay các khe hở nhỏ.[59]
Dưới nước
[sửa | sửa mã nguồn] Bài chi tiết: Rắn biển
Khi bơi, các sóng trở nên lớn hơn khi chúng di chuyển xuống phía dưới thân rắn, và tốc độ truyền sóng về phía sau nhanh hơn tốc độ con rắn di chuyển về phía trước.[60] Lực đẩy được sinh ra nhờ sự đẩy của thân rắn vào nước, thu được sự trượt được quan sát thấy. Mặc cho các điểm tương đồng tổng thể, các nghiên cứu chỉ ra rằng kiểu kích hoạt cơ trong môi trường nước là khác hẳn khi so với chuyển động sóng ngang trên cạn, và điều đó biện minh cho việc gọi chúng là các cách thức tách biệt.[61] Tất cả các loài rắn đều có thể bò theo kiểu lượn sóng ngang về phía trước (với các sóng di chuyển về phía sau), nhưng chỉ có rắn biển mới có kiểu di chuyển đảo ngược (di chuyển về phía sau với các sóng di chuyển về phía trước).
Uốn lượn nghiêng
[sửa | sửa mã nguồn] Xem thêm: Uốn lượn nghiêng
Kiểu di chuyển thường được các loài rắn trong siêu họ Colubroidea (các họ Colubridae, Elapidae và Viperidae) thực hiện khi phải di chuyển trong môi trường không có các điểm mấp mô để đẩy vào (trình diễn kiểu chuyển động sóng ngang là không thể), chẳng hạn như trên nền bùn trơn tuột hay trên cồn cát. Uốn lượn nghiêng là dạng biến đổi của chuyển động sóng ngang, trong đó tất cả các đoạn thân hướng theo cùng một hướng vẫn tiếp xúc với mặt nền trong khi các đoạn thân khác được nâng lên, tạo ra kiểu chuyển động "lăn ngang" kì dị.[62][63] Cách thức vận động này khắc phục được bản chất trơn trượt của cát hay bùn bằng cách tựa vào chỉ các phần tĩnh tại trên cơ thể, bằng cách này giảm thiểu sự trơn trượt.[62] Bản chất tĩnh tại của các tiếp điểm có thể được chỉ ra từ các dấu vết của con rắn di chuyển kiểu uốn lượn nghiêng, với vết hằn của mỗi vảy bụng là rõ nét, không bị nhòe. Phương thức vận động này có tiêu hao năng lượng rất thấp, thấp hơn ⅓ tiêu hao năng lượng cho một con thằn lằn chạy hay một con rắn di chuyển kiểu sóng ngang trên cùng một khoảng cách. Trái với niềm tin phổ biến, không có chứng cứ cho thấy chuyển động kiểu uốn lượn nghiêng là vì cát nóng.[62]
Đàn concertina
[sửa | sửa mã nguồn] Bài chi tiết: Chuyển động concertinaKhi không có điểm đẩy và không đủ không gian để sử dụng kiểu uốn lượn nghiêng do các hạn chế theo phương ngang, như bên trong đường ống, thì rắn dựa vào kiểu vận động concertina.[55][63] Trong cách thức này, rắn chống phần thân sau vào thành đường ống trong khi phần thân trước duỗi thẳng.[62] Phần thân trước sau đó uốn cong tạo thành một điểm tựa, và phần thân sau được duỗi thẳng ra và được kéo về phía trước. Cách thức vận động này chậm và rất tốn năng lượng, tới 7 lần so với chuyển động sóng ngang trên cùng một khoảng cách. Điều này là do sự dừng lại và bắt đầu được lặp đi lặp lại của các phần thân cũng như do sự cần thiết của việc sử dụng các cố gắng cơ bắp tích cực để chống vào thành đường ống.
Thẳng
[sửa | sửa mã nguồn] Bài chi tiết: Vận động thẳngCách thức vận động chậm nhất của rắn là vận động thẳng, cũng là cách thức duy nhất mà rắn không cần uốn cong cơ thể nó sang ngang, mặc dù nó có thể vẫn làm vậy khi gặp chỗ rẽ.[64] Trong cách thức này, các vảy bụng được nâng lên và được kéo về phía trước, trước khi cơ thể được hạ xuống và được kéo qua chúng. Các sóng di chuyển và ngưng trệ chuyển về phía sau, tạo ra một chuỗi các sóng gợn trên da.[64] Các xương sườn của rắn không di động trong phương thức vận động này và đây cũng là phương pháp được các loài trăn lớn trong các họ Pythonidae, Boidae và các loài rắn lục của họ Viperidae sử dụng thường xuyên nhất khi chúng lén đuổi theo con mồi ngang qua nền trống trải do chuyển động của rắn theo kiểu này là tinh tế và khó bị con mồi phát hiện.[62]
Khác
[sửa | sửa mã nguồn]Sự vận động của các loài rắn trong các môi trường sống trên cây mới chỉ được nghiên cứu gần đây.[65] Khi ở trên các cành cây, rắn sử dụng vài phương thức vận động, phụ thuộc vào kết cấu vỏ cây và tùy từng loài. Nói chung, rắn sẽ sử dụng dạng vận động concertina được biến đổi trên các cành cây nhẵn nhụi, nhưng sẽ sử dụng dạng chuyển động sóng ngang nếu có các tiếp điểm. Rắn di chuyển nhanh hơn trên các cành nhỏ và khi có các tiếp điểm, ngược lại với các động vật có chân, là những con vật di chuyển nhanh hơn trên các cành to với ít các 'mấu' hơn.
Các loài rắn cườm (Chrysopelea) ở Đông Nam Á phóng thân mình từ các đầu cành, trải rộng các xương sườn ra và chuyển động kiểu sóng ngang khi chúng lướt đi giữa các cây.[62][66][67] Các loài rắn này có thể thực hiện sự lướt đi có kiểm soát tới hàng chục mét, phụ thuộc vào cao độ phóng và thậm chí có thể xoay mình trong không trung.[62][66]
Sinh sản
[sửa | sửa mã nguồn]Mặc dù các loài rắn sử dụng một loạt các phương thức sinh sản, nhưng tất cả đều là thụ tinh trong. Điều này được thực hiện với sự hỗ trợ của một cơ quan có chạc đôi, gọi là bán dương vật, có thể thò ra thụt vào, nằm ở phần đuôi rắn đực.[68] Các bán dương vật thường xẻ khía, có móc hay có gai để kẹp vào vách lỗ huyệt của rắn cái.[68]
Phần lớn các loài rắn đẻ trứng và nói chung chúng sẽ bỏ rơi trứng ngay sau khi đẻ. Tuy nhiên, một số loài (như rắn hổ mang chúa) lại thực sự làm tổ và vẫn ở gần các con rắn con sau khi ấp.[68] Phần lớn các loài trăn họ Pythonidae cuộn tròn xung quanh ổ trứng của chúng và ở đó cho tới khi trứng nở.[69] Một con trăn cái sẽ không rời ổ trứng, ngoại trừ thỉnh thoảng phải bò ra tắm nắng hay uống nước. Nó thậm chí còn "rùng mình" để sinh nhiệt nhằm ấp trứng.[69]
Một số loài rắn là sinh noãn thai và giữ các quả trứng trong cơ thể cho tới khi chúng gần như đã sẵn sàng để nở.[70][71] Gần đây người ta đã xác nhận rằng một vài loài rắn là động vật đẻ con thật sự, như Boa constrictor và trăn anaconda xanh (Eunectes murinus), nuôi con non của chúng thông qua nhau thai cũng như túi noãn hoàng, và điều này là rất bất thường ở các loài bò sát, cũng như ở bất cứ loài động vật nào khác không phải là cá mập trong họ Carcharhinidae hay thú có nhau thai (Eutheria).[70][71] Sự cầm giữ lại trứng và sinh con non thường gắn với các môi trường lạnh hơn, cũng giống như sự cầm giữ con non trong cơ thể con mẹ.[68][71]
Tương tác với con người
[sửa | sửa mã nguồn]
Rắn cắn
[sửa | sửa mã nguồn] Bài chi tiết: Rắn cắn
Thông thường rắn không tấn công con người, trừ khi bị giật mình hay bị thương. Nói chung phần lớn các loài rắn cố gắng tránh tiếp xúc khi đối mặt với con người. Ngoại trừ các loài trăn lớn, các loài rắn không có nọc độc không phải là mối đe dọa đối với người. Những vết cắn của rắn không nọc thường là vô hại do răng của chúng không được thiết kế để xé rách hay gây ra những vết thương sâu, mà là để tóm giữ. Tuy vậy vẫn có khả năng xảy ra tổn thương mô và nhiễm trùng từ những vết do rắn không nọc cắn. Ngược lại, rắn có nọc độc (rắn độc) lại tiềm ẩn nguy hiểm lớn hơn nhiều cho con người,[6]:209 cho dù những ca tử vong do rắn cắn không phải là quá phổ biến. Những vết cắn không dẫn tới tử vong do rắn độc gây ra có thể vẫn dẫn tới hậu quả là phải cắt cụt một phần chân tay. Trong số khoảng 725 loài rắn có nọc độc trên toàn thế giới thì chỉ khoảng 250 loài có thể giết chết con người chỉ bằng một nhát cắn. Tại Australia trung bình chỉ một lần rắn cắn gây tử vong mỗi năm, nhưng tại Ấn Độ có tới 250.000 lần rắn cắn được ghi nhận trong năm, gây ra tới 50.000 vụ tử vong.[75]
Việc xử lý vết rắn cắn cũng thay đổi tùy theo từng loại vết cắn. Phương pháp thông dụng và hiệu quả nhất là thông qua huyết thanh kháng nọc rắn (antivenom hay antivenin), một loại huyết thanh được chế ra từ nọc rắn. Một số loại antivenom là chuyên biệt theo loài (đơn hóa trị, đặc hiệu) trong khi một số khác được chế ra để sử dụng cho nhiều loài (đa hóa trị). Chẳng hạn tại Hoa Kỳ, gần như tất cả các loài rắn độc đều là rắn vipe hốc lõm (phân họ Crotalinae trong họ Viperidae), ngoại lệ duy nhất là các loài rắn san hô (các chi Micrurus, Micruroides thuộc họ Elapidae). Để sản xuất antivenom, hỗn hợp nọc của các loài rắn khác nhau như rắn chuông (Crotalus, Sistrurus), rắn đầu đồng (Agkistrodon contortrix) và rắn miệng bông (Agkistrodon spp.) được tiêm vào cơ thể ngựa với liều lượng tăng dần cho tới khi ngựa được miễn dịch. Sau đó máu được chiết ra từ ngựa đã miễn dịch. Huyết thanh được tách ra để tinh chế tiếp (để loại bỏ các protein ngoại lai) và sấy thăng hoa. Nó được hoàn nguyên với nước cất và trở thành huyết thanh kháng nọc rắn. Vì lý do này mà những người bị dị ứng với ngựa rất dễ bị dị ứng với huyết thanh kháng nọc rắn.[76] Huyết thanh kháng nọc rắn cho những loài nguy hiểm hơn (như mamba (Dendroaspis), taipan (Oxyuranus) và rắn hổ mang (Naja)) được sản xuất theo phương pháp tương tự tại Ấn Độ, Nam Phi và Australia, mặc dù các loại huyết thanh kháng nọc rắn này là đặc hiệu theo loài.
Thôi miên rắn
[sửa | sửa mã nguồn] Bài chi tiết: Thôi miên rắn
Tại một vài nơi trên thế giới, đặc biệt tại Ấn Độ, thôi miên rắn là cuộc trình diễn ven đường do những người làm nghề thôi miên rắn trình diễn. Trong những cuộc trình diễn như vậy, người thôi miên rắn mang theo một chiếc giỏ chứa một con rắn để thực hiện việc dường như là ông ta thôi miên con rắn bằng cách thổi một số điệu nhạc du dương bằng một nhạc cụ dạng sáo để con rắn phản ứng theo.[77][77][78] Sự thật thôi miên rắn là lừa đảo và tàn nhẫn với động vật, loài rắn không có tai ngoài và chẳng hề nghe thấy tiếng nhạc. Việc rắn phồng mang lên và đung đưa theo điệu nhạc chẳng qua vì nó coi chiếc kèn pungi là mối đe doạ và đang đứng lên trong tư thế phòng thủ. Nếu quan sát kĩ hơn, có thể thấy con rắn hổ mang chỉ nhìn chằm chằm vào nhạc cụ của các thầy "phủ thủy" – đặc biệt là chuyển động của ngón tay xung quanh chiếc kèn.
Thật ra rắn đã bị nhiều 'thầy' nhổ răng. Ngoài ra, để đảm bảo an toàn, những thầy "phù thủy" sẽ nhổ răng nanh hoặc khâu kín miệng rắn. Đιềυ này vừa khiến việc thôi miên không gặp trở ngại vừa khiến họ thu được thêm lợi nhuận thông qua việc cho du khách tạo dáng chụp ảnh. Một con rắn bị nhổ răng. Số phận của những chú rắn không tốt đẹp như bạn tưởng tượng. Вị nhổ hết răng nanh, chúng sẽ mất đi khả năng ăn uống và chỉ sống được tối đa 2 tháng.[79]
Luật bảo vệ động vật hoang dã năm 1972 của Ấn Độ về mặt kỹ thuật cấm thôi miên rắn trên cơ sở làm giảm sự đối xử độc ác với động vật. Những người thôi miên rắn khác còn trình diễn cuộc đấu giữa rắn với cầy mangut, trong đó cả hai con vật có một cuộc đấu giả vờ; tuy nhiên điều này là không phổ biến do cả rắn lẫn cầy mangut đều có thể bị thương nặng hay tử vong. Nghề thôi miên rắn tại Ấn Độ đang tàn lụi do sự cạnh tranh từ các dạng giải trí hiện đại cũng như do luật môi trường ngăn cấm việc hành nghề này.[77]
Bắt rắn
[sửa | sửa mã nguồn]Bộ lạc Irulas ở Andhra Pradesh và Tamil Nadu thuộc Ấn Độ hành nghề bắt rắn trong nhiều thế hệ. Họ có nhiều kiến thức và kinh nghiệm về rắn trên thực địa. Nói chung họ bắt rắn với sự hỗ trợ của một cây sào. Trước đây, người Irulas đã bắt hàng nghìn con rắn để lấy da. Sau khi ngành công nghiệp da rắn bị cấm triệt để và sự bảo vệ rắn theo Luật bảo vệ động vật hoang dã năm 1972 tại Ấn Độ thì họ đã lập ra Hội hợp tác bắt rắn Irula và chuyển sang bắt rắn để lấy nọc và thả chúng ra sau 4 lần lấy nọc. Nọc rắn thu thập theo cách này được dùng để sản xuất huyết thanh kháng nọc rắn cũng như phục vụ cho nghiên cứu y sinh học và sản xuất các dược phẩm khác.[80]
Bên cạnh đó còn có những người bắt rắn chuyên nghiệp khác. Hành nghề bắt rắn ngày nay còn có các nhà bò sát học với công cụ là các cây sào dài với đầu sào hình chữ V. Một vài người dẫn chương trình truyền hình về động vật hoang dã, như Bill Haast, Austin Stevens, Steve Irwin, Jeff Corwin v.v. còn bắt rắn trực tiếp bằng tay không.
Tiêu thụ
[sửa | sửa mã nguồn] Bài chi tiết: Thịt rắn

Trong khi đa phần các nền văn hóa không dùng rắn làm thực phẩm thì tại một số quốc gia việc sử dụng rắn làm thực phẩm lại được chấp nhận hay thậm chí còn được coi là đặc sản, do các thức ăn chế biến từ rắn được đánh giá cao về tác dụng y học. Món xúp rắn trong ẩm thực Quảng Đông được người dân tại đây dùng trong mùa thu do họ coi nó có tác dụng làm ấm cơ thể. Trong các nền văn hóa phương Tây thì việc ăn thịt rắn cũng từng xảy ra khi gặp phải những năm đói kém.[81] Việc ăn thịt rắn chuông đã nấu chín là một ngoại lệ, với việc tiêu thụ nói chung phổ biến tại các khu vực ở miền trung tây nước Mỹ. Tại các quốc gia châu Á, như Trung Quốc, Đài Loan, Thái Lan, Indonesia, Việt Nam và Campuchia, việc uống rượu pha tiết rắn—cụ thể là rắn hổ mang—được cho là làm gia tăng khả năng tình dục.[82] Việc lấy tiết rắn được thực hiện khi con rắn còn sống và nó được pha với một vài loại rượu mạnh hay rượu mùi để cải thiện mùi vị.[82]
Tại một số quốc gia châu Á việc sử dụng rắn ngâm trong rượu cũng được chấp nhận. Trong trường hợp này nguyên cả con rắn hay nguyên cả con của một vài loại rắn được ngâm trong bình rượu mạnh hay rượu mùi. Người ta cho rằng rượu rắn có tác dụng tốt với cơ thể (và rượu rắn cũng được bán đắt hơn). Một ví dụ là loài rắn lục Habu (Trimeresurus flavoviridis) đôi khi được ngâm trong rượu Awamori của người dân Okinawa và được gọi là "Habu Sake".[83]
Các học viên của lực lượng đặc nhiệm Quân đội Hoa Kỳ cũng được huấn luyện bắt, giết và ăn thịt rắn để có thể sống sót trong các hoàn cảnh đặc biệt; và điều này đã làm người ta gán cho họ tên hiệu là "snake eaters" (những kẻ ăn rắn).
Rượu rắn (蛇酒; xà tửu) là loại rượu ngâm nguyên cả con rắn trong các loại rượu sản xuất từ gạo hay ngũ cốc. Việc uống rượu rắn tại Trung Quốc lần đầu tiên được ghi chép lại có từ thời Tây Chu và được coi là có tác dụng chữa bệnh và làm cường tráng cơ thể theo y học cổ truyền Trung Hoa.[84]
Động vật cảnh
[sửa | sửa mã nguồn]Tại phương Tây, một vài loài rắn (đặc biệt là các loài dễ thuần hóa như trăn hoàng gia (Python regius) và rắn săn chuột đỏ (Pantherophis guttatus) được nuôi làm động vật cảnh. Để đáp ứng nhu cầu này một ngành công nghiệp nuôi nhốt động vật bò sát đã phát triển. Các loại rắn sinh đẻ nuôi nhốt có xu hướng dễ nuôi làm động vật cảnh hơn so với các loại rắn bắt được trong tự nhiên.[85] Rắn cảnh dễ nuôi hơn so với nhiều loại động vật cảnh truyền thống khác do chúng không cần nhiều không gian (do phần lớn rắn cảnh không dài quá 1,5 m và chúng cũng không cần phải cho ăn uống quá thường xuyên (thường chỉ một lần trong vòng 5-14 ngày). Một vài loại rắn cảnh có thể sống trên 40 năm nếu được chăm sóc chu đáo.
Văn hóa
[sửa | sửa mã nguồn]Biểu tượng
[sửa | sửa mã nguồn] Bài chi tiết: Rắn (biểu tượng)
Trong lịch sử Ai Cập, rắn có một vai trò quan trọng với rắn hổ mang sông Nil trang điểm cho vương miện của các pharaoh trong thời kỳ cổ đại. Nó được tôn thờ như là một vị thần và từng được sử dụng cho các mục đích độc ác: nó là kẻ sát nhân trong một vụ tự sát theo nghi lễ của Cleopatra.
Trong thần thoại Hy Lạp rắn thường gắn với các địch thủ nguy hiểm chết người, nhưng điều này không có nghĩa rằng rắn là biểu tượng của điều ác độc; trên thực tế rắn là biểu tượng gắn liền với đất hay âm phủ. Con rắn chín đầu Lernaean Hydra mà Hercules đánh bại và ba chị em Gorgon đều là con của Gaia, nữ thần đất.[86] Medusa là một trong số ba chị em Gorgon mà Perseus đã đánh bại.[86] Medusa được mô tả là một vị thần bất tử gớm ghiếc, với mái tóc là những con rắn và có phép thuật biến những kẻ đàn ông thành đá chỉ bằng ánh mắt.[86] Sau khi giết chết Medusa, Perseus đã dâng đầu nàng cho Athena để gắn vào chiếc khiên che ngực gọi là Aegis.[86] Các Titan cũng được mô tả là có các con rắn thay vì có chân vì cùng một lý do—họ đều là con của Gaia với Ouranos (Uranus), vì thế họ cũng gắn liền với đất.

Truyền thuyết về sự ra đời của Thebes đề cập tới một con rắn quái vật khổng lồ ngăn giữ suối nước mà dân cư của khu định cư này lấy nước. Trong cuộc chiến để giết chết con rắn, những người bạn đồng hành của nhà sáng lập khu định cư là Cadmus đều bị giết chết – điều này dẫn tới thuật ngữ "chiến thắng kiểu Cadmus" (nghĩa là chiến thắng với sự tự hủy diệt bản thân).
Ba biểu tượng y học liên quan tới rắn còn được sử dụng cho tới ngày nay là Chén Hygieia, biểu tượng cho dược học, và Caduceus cùng Gậy Asclepius là biểu tượng cho y tế nói chung.[42]
Ấn Độ thường được gọi là vùng đất rắn và huyền ảo trong các tập tục liên quan tới rắn.[87] Ngay cả ngày nay rắn vẫn còn được thờ phụng với nhiều phụ nữ rót sữa vào các hốc rắn (mặc cho ác cảm của rắn đối với sữa).[87] Rắn hổ mang có trên cổ thần Shiva còn thần Vishnu thường được vẽ trong tư thế đang ngủ trên con rắn bảy đầu hoặc với các cuộn rắn.[88] Có một số đền miếu tại Ấn Độ chỉ dành cho rắn hổ mang, đôi khi được gọi là Nagraj (vua rắn) và người ta tin rằng rắn là biểu tượng của sự trù phú. Có một lễ hội Hindu gọi là Nag Panchami mỗi năm (tổ chức vào ngày thứ năm sau ngày Amavasya của tháng Shraavana), trong ngày đó những con rắn được tôn thờ và vái lạy. Xem thêm Nāga.
Tại Ấn Độ còn có truyền thuyết khác về rắn. Nói chung được biết đến như là những con rắn "Ichchhadhari" trong tiếng Hindi. Những con rắn này có thể có hình dáng của bất kỳ sinh vật nào, nhưng thích biến hình thành dạng người. Những con rắn thần bí này chiếm giữ viên ngọc quý gọi là "Mani" quý báu hơn cả kim cương. Có nhiều câu chuyện cổ tích tại Ấn Độ kể về những kẻ tham lam cố đoạt viên ngọc quý này và đều có chung một kết cục là bị giết chết.
Ouroboros là biểu tượng gắn với nhiều tôn giáo và tập quán khác nhau, và được coi là có liên quan tới giả kim thuật. Ouroboros hay Oroboros là con rắn hay con rồng cắn đuôi của chính nó theo chiều kim đồng hồ (tính từ đầu tới đuôi) vẽ thành vòng tròn, tượng trưng cho chu trình sống, chết và tái sinh, dẫn tới sự bất tử.
Tị là một trong số 12 địa chi của Thiên Can Địa Chi, trong lịch Trung Quốc.
Nhiều nền văn hóa Peru cổ đại thờ phụng thiên nhiên.[89] Người dân của các nền văn hóa này đề cao các con vật và thường vẽ các con rắn trong các bức họa của mình.[90]
Các nhân vật phim hoạt hình
[sửa | sửa mã nguồn]- Trăn Ấn Độ Kaa (The Jungle Book)
- Rắn lục xanh Master Viper (Kungfu Panda 1-2-3)
- Trăn xiết mồi Savio (Penguins Of Madagascar)
- Rắn hổ mang Ai Cập Ushari (The Lion Guard)
- Phù thủy Jafar hòa thành rắn hổ mang (Aladdin) (1992)
Sử dụng trong show
[sửa | sửa mã nguồn]Trăn và rắn lục được sử dụng làm xiếc và các game show đòi hỏi sự gan dạ.
Xem thêm
[sửa | sửa mã nguồn]- Truyền thuyết Bạch Xà
- Rắn độc
Ghi chú
[sửa | sửa mã nguồn]- ^ Hsiang, A. Y.; Field, D. J.; Webster, T. H.; Behlke, A. D.; Davis, M. B.; Racicot, R. A.; Gauthier, J. A. (2015). "The origin of snakes: Revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record". BMC Evolutionary Biology. Quyển 15. doi:10.1186/s12862-015-0358-5. ISSN 1471-2148.{{Chú thích tạp chí}}: Quản lý CS1: DOI truy cập mở nhưng không được đánh ký hiệu (liên kết)
- ^ a b c d e f g h i j k l Serpentes (TSN 174118) tại Hệ thống Thông tin Phân loại Tích hợp (ITIS). Truy cập ngày 20-8 năm 2007.
- ^ a b Bauchot, Roland (1994). Snakes: A Natural History. New York City, NY, Hoa Kỳ: Sterling Publishing Co., Inc. tr. 220. ISBN 1-4027-3181-7.
- ^ a b c Danh sách các loài rắn tại Reptile Database. Tra cứu 07-08-2012.
- ^ Murphy; Henderson, JC; RW (1997). Tales of Giant Snakes: A Historical Natural History of Anacondas and Pythons. Florida, Hoa Kỳ: Krieger Pub. Co. tr. 221. ISBN 0-89464-995-7.{{Chú thích sách}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- ^ a b c d e f g h i j k l m n o p q Mehrtens, JM (1987). Living Snakes of the World in Color. New York City, NY, Hoa Kỳ: Sterling Publishers. tr. 480. ISBN 0-8069-6460-X.
- ^ Proto-IE: *(s)nēg-o-, Nghĩa là: rắn, Tiếng Ấn cổ: nāgá- m. 'rắn', tiếng German: *snēk-a- m., *snak-an- m., *snak-ō f.; *snak-a- vb., tiếng Nga. nghĩa là: жаба (змея), Dẫn nguồn: WP (Vergleichendes Wörterbuch der indogermanischen Sprachen) II 697 f.
- ^ "Definition of serpent". Merriam-Webster Online Dictionary. Lưu trữ bản gốc ngày 17 tháng 10 năm 2007. Truy cập ngày 12 tháng 10 năm 2006.
- ^ a b Michael S. Y. Lee & Andrew F. Hugall, Robin Lawson & John D. Scanlon (2007). "Phylogeny of snakes (Serpentes): combining morphological and molecular data in likelihood, Bayesian and parsimony analyses". Systematics and Biodiversity. Quyển 5 số 4. tr. 371–389. doi:10.1017/S1477200007002290.
- ^ Vidal N., Marin J., Morini M., Donnellan S., Branch W. R., Thomas R., Vences M., Wynn A., Cruaud C. & Hedges S. B., 2010: Blindsnake evolutionary tree reveals long history on Gondwana. Biology Letters, 6 (4): 558-561, doi:10.1098/rsbl.2010.0220
- ^ Durand, J.F. (2004). "The origin of snakes". Geoscience Africa 2004. Abstract Volume, Đại học Witwatersrand, Johannesburg, Nam Phi, tr. 187.
- ^ Vidal N., Rage J.-C., Couloux A. và Hedges S.B. (2009). "Snakes (Serpentes)". Tr. 390-397 trong Hedges S.B. và Kumar S. (chủ biên), The Timetree of Life. Nhà in Đại học Oxford.
- ^ Mehrtens JM. 1987. Living Snakes of the World in Color. New York: Sterling Publishers. 480 tr. ISBN 0-8069-6460-X.
- ^ a b c d e Sanchez, Alejandro. "Diapsids III: Snakes". Father Sanchez's Web Site of West Indian Natural History. Truy cập ngày 7 tháng 8 năm 2012.
- ^ a b "New Fossil Snake With Legs". UNEP WCMC Database. Washington, D.C.: American Association For The Advancement Of Science. Bản gốc lưu trữ ngày 25 tháng 12 năm 2007. Truy cập ngày 7 tháng 8 năm 2012.
- ^ a b Mc Dowell, Samuel (1972). "The evolution of the tongue of snakes and its bearing on snake origins". Evolutionary Biology. Quyển 6. tr. 191–273.
- ^ Apesteguía, Sebastián; Zaher, Hussam (2006). "A Cretaceous terrestrial snake with robust hindlimbs and a sacrum". Nature. Quyển 440 số 7087. tr. 1037–1040. doi:10.1038/nature04413. PMID 16625194. Truy cập ngày 7 tháng 8 năm 2012.
- ^ a b c Mertens, Robert (1961). "Lanthanotus: an important lizard in evolution". Sarawak Museum Journal. Quyển 10. tr. 320–322.
- ^ a b Conant R, Collins JT. 1991. A Field Guide to Reptiles and Amphibians: Eastern and Central North America. Houghton Mifflin, Boston. 450 tr. 48 tiêu bản. ISBN 0-395-37022-1.
- ^ Natural History Information Centre, Auckland War Memorial Museum. "Natural History Questions". Auckland War Memorial Museum | Tamaki Paenga Hira. Auckland, New Zealand: Auckland War Memorial Museum. Q. Are there any snakes in New Zealand?. Bản gốc lưu trữ ngày 12 tháng 7 năm 2012. Truy cập ngày 26 tháng 4 năm 2012.
- ^ Pough và ctv. (2002) [1992]. Herpetology: Third Edition. Pearson Prentice Hall. ISBN 0-13-100849-8.
- ^ a b McDiarmid RW, Campbell JA, Touré T. 1999. Snake Species of the World: A Taxonomic and Geographic Reference, vol. 1. Herpetologists' League. 511 tr. ISBN 1-893777-00-6 (series). ISBN 1-893777-01-4 (volume).
- ^ Spawls S, Branch B. 1995. The Dangerous Snakes of Africa. Ralph Curtis Books. Dubai: Oriental Press. 192 tr. ISBN 0-88359-029-8.
- ^ Parker HW, Grandison AGC. 1977. Snakes -- a natural history. Ấn bản lần 2. Bảo tàng Anh (Lịch sử Tự nhiên) và Nhà in Đại họcCornell. 108 tr. 16 tiêu bản. LCCCN 76-54625. ISBN 0-8014-1095-9 (vải), ISBN 0-8014-9164-9 (giấy).
- ^ Spawls S, Howell K, Drewes R, Ashe J. 2004. A Field Guide To The Reptiles Of East Africa. Luân Đôn: A & C Black Publishers Ltd. 543 tr. ISBN 0-7136-6817-2.
- ^ Elapidae tại Reptarium.cz Reptile Database. Tra cứu 3-12 2008.
- ^ Harline, P H (1971). "Physiological basis for detection of sound and vibration in snakes" (pdf). J. Exp. Biol. Quyển 54 số 2. tr. 349–371.
- ^ "Auditory Localization of Ground-Borne Vibrations in Snakes". Phys. Rev. Lett. Quyển 100. 2008. tr. 048701.
- ^ Lisa Zyga (ngày 13 tháng 2 năm 2008). "Desert Snake Hears Mouse Footsteps with its Jaw". PhysOrg.
- ^ Cogger, H 1993 Fauna of Australia. Vol. 2A Amphibia and Reptilia. Australian Biological Resources Studies, Canberra.
- ^ Arnold, E.N. (1984). "Evolutionary aspects of tail shedding in lizards and their relatives". Journal of Natural History. Quyển 18 số 1. tr. 127–169. doi:10.1080/00222938400770131.
- ^ N. B. Ananjeva và N. L. Orlov (1994) Caudal autotomy in Colubrid snake Xenochrophis piscator from Vietnam. Russian Journal of Herpetology 1(2)
- ^ a b c d e f Mader, Douglas (tháng 6 năm 1995). "Reptilian Anatomy". Reptiles. Quyển 3 số 2. tr. 84–93.
- ^ "CTV: Ancient, gargantuan snakes ate crocs for breakfast". Bản gốc lưu trữ ngày 6 tháng 2 năm 2009. Truy cập ngày 20 tháng 2 năm 2013.
- ^ Jesús Antonio Rivas (2000). The life history of the green anaconda (Eunectes murinus), with emphasis on its reproductive Biology (pdf) (Luận văn). Đại học Tennessee. Lưu trữ ngày 3 tháng 3 năm 2016 tại Wayback Machine
- ^ S. Blair Hedges (ngày 4 tháng 8 năm 2008). "At the lower size limit in snakes: two new species of threadsnakes (Squamata: Leptotyphlopidae: Leptotyphlops) from the Lesser Antilles" (pdf). Zootaxa. Quyển 1841. tr. 1–30. Truy cập ngày 7 tháng 8 năm 2012.
- ^ Boback S. M.; Guyer C. (2003). "Empirical Evidence for an Optimal Body Size in Snakes". Evolution 57 (2): 345. doi:10.1554/0014-3820(2003)057[0345:EEFAOB]2.0.CO;2. ISSN 0014-3820. PMID 12683530.
- ^ Smith Malcolm A. The Fauna of British India, Including Ceylon and Burma. Vol I, Loricata and Testudines. tr. 30.
- ^ a b c d Are snakes slimy? Lưu trữ ngày 3 tháng 5 năm 2006 tại Wayback Machine tại Singapore Zoological Garden's Docent. Tra cứu 07-08-2012.
- ^ Part III: Scales of Lizards and Snakes tại WhoZoo. Tra cứu 7-8-2012.
- ^ General Snake Information Lưu trữ ngày 31 tháng 3 năm 2009 tại Wayback Machine tại South Dakota Game, Fish and Parks. Tra cứu 7-8-2012.
- ^ a b Wilcox, Robert A; Whitham, Emma M (ngày 15 tháng 4 năm 2003). "The symbol of modern medicine: why one snake is more than two". Annals of Internal Medicine. Quyển 138 số 8. tr. 673–7. PMID 12693891. Truy cập ngày 7 tháng 8 năm 2012.
- ^ Reptile Senses: Understanding Their World Lưu trữ ngày 19 tháng 2 năm 2015 tại Wayback Machine.
- ^ a b c d e Cogger(1991), tr. 180.
- ^ a b c d e Freiberg (1984), tr. 125.
- ^ a b Freiberg (1984), tr. 123.
- ^ a b c d Freiberg (1984), tr. 126.
- ^ Fry, Brian G.; Vidal, Nicholas; Norman, Janette A.; Vonk, Freek J.; Scheib, Holger; Ramjan, S. F. Ryan; Kuruppu, Sanjaya; Fung, Kim Y. C.; Hedges, S. Blair; Richardson, Michael K.; Hodgson, Wayne C.; Vera, Ignjatovic; Robyn, Summerhayes; Elazar, Kochva (2006). "Early evolution of the venom system in lizards and snakes". Nature (Letters). Quyển 439 số 7076. tr. 584–588. doi:10.1038/nature04328. PMID 16292255.
- ^ a b c d Behler (1979) tr. 581.
- ^ Bebler (1979) tr. 581.
- ^ Hori, Michio; Asami, Takahiro; Hoso, Masaki (2007). "Right-handed snakes: convergent evolution of asymmetry for functional specialization". Biology Letters. Quyển 3 số 2. tr. 169–72. doi:10.1098/rsbl.2006.0600. PMC 2375934. PMID 17307721.
- ^ Freiberg (1984), tr. 125–127.
- ^ Rosenfeld (1989), tr. 11.
- ^ Tattersall, GJ; Milsom, WK; Abe, AS; Brito, SP; Andrade, DV (2004). "The thermogenesis of digestion in rattlesnakes". Journal of Experimental Biology. Quyển 207 số Pt 4. The Company of Biologists. tr. 579–585. doi:10.1242/jeb.00790. PMID 14718501. Truy cập ngày 26 tháng 5 năm 2006.
- ^ a b c d e Cogger(1991), tr. 175.
- ^ a b Gray, J. (1946). "The mechanism of locomotion in snakes". Journal of experimental biology. Quyển 23 số 2. tr. 101–120. PMID 20281580.
- ^ a b Hekrotte, Carlton (1967). "Relations of Body Temperature, Size, and Crawling Speed of the Common Garter Snake, Thamnophis s. sirtalis". Copeia. Quyển 23 số 4. tr. 759–763. doi:10.2307/1441886. JSTOR 1441886.
- ^ Walton, M.; Jayne, B.C.; Bennett, A.F. (1967). "The energetic cost of limbless locomotion". Science. Quyển 249 số 4968. tr. 524–527. doi:10.1126/science.249.4968.524. PMID 17735283.
- ^ a b Gray, J; H.W., H (1950). "Kinetics of locomotion of the grass snake". Journal of experimental biology. Quyển 26 số 4. tr. 354–367.
- ^ Gray, J; Lissman (1953). "Undulatory propulsion". Quarterly Journal of Microscopical Science. Quyển 94. tr. 551–578.
- ^ Jayne, B.C. (1988). "Muscular mechanisms of snake locomotion: an electromyographic study of lateral undulation of the Florida banded water snake (Nerodia fasciata) and the yellow rat snake (Elaphe obsoleta)". Journal of Morphology. Quyển 197 số 2. tr. 159–181. doi:10.1002/jmor.1051970204. PMID 3184194.
- ^ a b c d e f g Cogger(1991), tr. 177.
- ^ a b Jayne, B.C. (1986). "Kinematics of terrestrial snake locomotion". Copeia. Quyển 1986 số 4. tr. 915–927. doi:10.2307/1445288. JSTOR 1445288.
- ^ a b Cogger (1991), tr. 176.
- ^ Astley, H.C.; Jayne, B.C. (2007). "Effects of perch diameter and incline on the kinematics, performance and modes of arboreal locomotion of corn snakes (Elaphe guttata)". Journal of Experimental Biology. Quyển 210 số Pt 21. tr. 3862–3872. doi:10.1242/jeb.009050. PMID 17951427.
- ^ a b Freiberg (1984), p. 135.
- ^ Socha, JJ (2002). "Gliding flight in the paradise tree snake". Nature. Quyển 418 số 6898. tr. 603–604. doi:10.1038/418603a. PMID 12167849.
- ^ a b c d Capula (1989), tr. 117.
- ^ a b Cogger (1991), tr. 186.
- ^ a b Capula (1989), tr. 118.
- ^ a b c Cogger (1991), tr. 182.
- ^ a b MedlinePlus > Snake bites Từ Tintinalli JE, Kelen GD, Stapcynski JS (chủ biên). Emergency Medicine: A Comprehensive Study Guide. Ấn bản lần 6. New York, NY: McGraw Hill; 2004. Cập nhật: 27/2/2008. Bởi: Stephen C. Acosta, MD, Department of Emergency Medicine, Portland VA Medical Center, Portland, OR. Do VeriMed Healthcare Network và David Zieve, MD, MHA, Medical Director, A.D.A.M., Inc. thẩm định. Tra cứu 9-3-2010.
- ^ Health-care-clinic.org > Snake Bite First Aid - Snakebite. Tra cứu 9-3-2010.
- ^ Ví dụ về hình ảnh vết rắn cắn tại MDconsult > Patient Education > Wounds, Cuts and Punctures, First Aid for.
- ^ Sinha, Kounteya (ngày 25 tháng 7 năm 2006). "No more the land of snake charmers..." The Times of India.
- ^ NCBI.nlm.nih.gov
- ^ a b c Bagla, Pallava (ngày 23 tháng 4 năm 2002). "India's Snake Charmers Fade, Blaming Eco-Laws, TV". National Geographic News. Truy cập ngày 26 tháng 11 năm 2007.
- ^ International Wildlife encyclopedia, ấn bản lần 3, tr. 482
- ^ "Sự thật tàn nhẫn sau thuật 'thôi miên rắn' của thầy phù thủy Ấn Độ". PHẬT GIÁO. Truy cập ngày 25 tháng 7 năm 2022.
- ^ Whitaker, Romulus & Captain, Ashok. Snakes of India: The Field Guide.(2004) tr. 11-13.
- ^ Irvine, F. R. (1954). "Snakes as food for man". British Journal of Herpetology. Quyển 1 số 10. tr. 183–189.
- ^ a b Flynn, Eugene (ngày 23 tháng 4 năm 2002). "Flynn Of The Orient Meets The Cobra". Fabulous Travel. Truy cập ngày 7 tháng 8 năm 2012.
- ^ Allen, David (ngày 22 tháng 7 năm 2001). "Okinawa's potent habu sake packs healthy punch, poisonous snake". Stars and Stripes. Bản gốc lưu trữ ngày 28 tháng 11 năm 2007. Truy cập ngày 26 tháng 11 năm 2007.
- ^ "蛇酒的泡制与药用(Sản xuất và tác dụng y học của rượu rắn)". ngày 9 tháng 4 năm 2007. Bản gốc lưu trữ ngày 6 tháng 7 năm 2011. Truy cập ngày 20 tháng 2 năm 2013.
- ^ Carl Ernest & George R. Zug, Molly Dwyer Griffin (1996). Snakes in Question: The Smithsonian Answer Book. Washington, DC: Smithsonian Books. tr. 203. ISBN 1-56098-648-4.
- ^ a b c d Bullfinch (2000) tr. 85.
- ^ a b Deane (1833). tr. 61.
- ^ Deane (1833). tr. 62–64.
- ^ Benson, Elizabeth (1972). The Mochica: A Culture of Peru. Luân Đôn: Thames and Hudson. ISBN 0-500-72001-0.
- ^ Berrin, Katherine; Larco Museum (1997). The Spirit of Ancient Peru: Treasures from the Museo Arqueológico Rafael Larco Herrera. New York: Thames and Hudson. ISBN 978-0-500-01802-6.
Đọc thêm
[sửa | sửa mã nguồn]- Behler, John L.; King, F. Wayne (1979). The Audubon Society Field Guide to Reptiles and Amphibians of North America. New York: Alfred A. Knopf. tr. 581. ISBN 0-394-50824-6.
- Bullfinch, Thomas (2000). Bullfinch's Complete Mythology. Luân Đôn: Chancellor Press. tr. 679. ISBN 0-7537-0381-5. Bản gốc lưu trữ ngày 9 tháng 2 năm 2009. Truy cập ngày 20 tháng 2 năm 2013.
- Capula, Massimo (1989). Simon & Schuster's Guide to Reptiles and Amphibians of the World. Behler. New York: Simon & Schuster. ISBN 0-671-69098-1.
- Coborn, John (1991). The Atlas of Snakes of the World. New Jersey: TFH Publications. ISBN 978-0-86622-749-0.
- Cogger, Harold; Zweifel, Richard (1992). Reptiles & Amphibians. Sydney: Weldon Owen. ISBN 0-8317-2786-1.
- Conant, Roger; Collins, Joseph (1991). A Field Guide to Reptiles and Amphibians Eastern/Central North America. Boston, Massachusetts: Houghton Mifflin Company. ISBN 0-395-58389-6.
- Deane, John (1833). The Worship of the Serpent. Whitefish, Montana: Kessinger Publishing. tr. 412. ISBN 1-56459-898-5.
- Ditmars, Raymond L (1906). Poisonous Snakes of the United States: How to Distinguish Them. New York: E. R. Sanborn. tr. 11.
- Ditmars, Raymond L (1931). Snakes of the World. New York: Macmillan. tr. 11. ISBN 978-0-02-531730-7.
- Ditmars, Raymond L (1933). Reptiles of the World: The Crocodilians, Lizards, Snakes, Turtles and Tortoises of the Eastern and Western Hemispheres. New York: Macmillan. tr. 321.
- Ditmars, Raymond L (1935). Snake-Hunters' Holiday. W. Bridges. New York: D. Appleton and Company. tr. 309.
- Ditmars, Raymond L (1939). A Field Book of North American Snakes. Garden City, New York: Doubleday, Doran & Co. tr. 305.
- Freiberg, Dr. Marcos; Walls, Jerry (1984). The World of Venomous Animals. New Jersey: TFH Publications. ISBN 0-87666-567-9.
- Gibbons, J. Whitfield; Gibbons, Whit (1983). Their Blood Runs Cold: Adventures With Reptiles and Amphibians. Alabama: Nhà in Đại học Alabama. tr. 164. ISBN 978-0-8173-0135-4.
- Mattison, Chris (2007). The New Encyclopedia of Snakes. New Jersey: Nhà in Đại học Princeton. tr. 272. ISBN 978-0-691-13295-2.
- McDiarmid, RW; Campbell, JA; Touré, T (1999). Snake Species of the World: A Taxonomic and Geographic Reference. Quyển 1. Herpetologists' League. tr. 511. ISBN 1-893777-00-6.
- Mehrtens, John (1987). Living Snakes of the World in Color. New York: Sterling. ISBN 0-8069-6461-8.
- Nóbrega Alves, RôMulo Romeu; Silva Vieira, Washington Luiz; Santana, Gindomar Gomes (2008). "Reptiles used in traditional folk medicine: conservation implications". Biodiversity and Conservation. Quyển 17 số 8. tr. 2037–2049. doi:10.1007/s10531-007-9305-0. Bản gốc lưu trữ ngày 17 tháng 4 năm 2003. Truy cập ngày 22 tháng 1 năm 2009.
- Romulus Whitaker (bản tiếng Anh); O.Henry Francis dịch từ tiếng Tamil (1996). நம்மை சுட்ரியுள்ள பாம்புகள் (Rắn xung quanh chúng ta, người Tamil). National Book Trust. ISBN 81-237-1905-1.{{Chú thích sách}}: Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- Rosenfeld, Arthur (1989). Exotic Pets. New York: Simon & Schuster. tr. 293. ISBN 978-0-671-47654-0.
- Spawls, Steven; Branch, Bill (1995). The Dangerous Snakes of Africa. đảo Sanibel, Florida: Ralph Curtis Publishing. tr. 192. ISBN 0-88359-029-8.
Liên kết ngoài
[sửa | sửa mã nguồn] Tư liệu liên quan tới Serpentes tại Wikimedia Commons
Tư liệu liên quan tới Serpentes tại Wikimedia Commons- Snake tại Encyclopædia Britannica (bằng tiếng Anh)
- Rắn tại Từ điển bách khoa Việt Nam
- "Worldwide Snake Species List". Snake Track.com.
- "Bibliography for "Serpentes"". Biodiversity Heritage Library.
- Rắn tại Encyclopedia of Life
- Rắn tại trang Trung tâm Thông tin Công nghệ sinh học quốc gia Hoa Kỳ (NCBI).
- "US Snakes". eNature. Bản gốc lưu trữ ngày 15 tháng 3 năm 2008. Truy cập ngày 20 tháng 2 năm 2013.
- "Snakes of the Indian Subcontinent". Naturemagics Kerala Photo Gallery. Bản gốc lưu trữ ngày 21 tháng 3 năm 2009. Truy cập ngày 20 tháng 2 năm 2013.
- "About Snake Cell". Snake Cell. (Ấn Độ)
- "Snake World". ReptilesWeb.com. Bản gốc lưu trữ ngày 3 tháng 7 năm 2008. Truy cập ngày 20 tháng 2 năm 2013.
- "What's That Snake?". OPLIN.
- "Herpetology Database". Swedish Museum of Natural History.
- BBC Nature: Lưu trữ ngày 1 tháng 3 năm 2014 tại Wayback Machine Snake news, and video clips from BBC programmes past and present.
- Rắn đẻ con hay đẻ trứng Lưu trữ ngày 3 tháng 8 năm 2020 tại Wayback Machine
| ||
|---|---|---|
| Ngành Chordata (Động vật có dây sống) • Lớp Reptilia (Động vật bò sát) • Bộ Squamata (Bò sát có vảy) • Phân bộ Serpentes (Rắn) | ||
| Cận bộ Alethinophidia | Acrochordidae • Aniliidae • Anomochilidae • Atractaspididae • Boidae • Bolyeriidae • Colubridae • Cylindrophiidae • Elapidae • Loxocemidae • Pythonidae • Tropidophiidae • Uropeltidae • Viperidae • Xenopeltidae | |
| Cận bộ Scolecophidia | Anomalepididae • Leptotyphlopidae • Typhlopidae • Gerrhopilidae • Xenotyphlopidae | |
| ||
|---|---|---|
| Khủng long |
|  |
| Rắn |
| |
| Khác |
| |
| Thẻ nhận dạng đơn vị phân loại |
|
|---|
| Cơ sở dữ liệu tiêu đề chuẩn | |
|---|---|
| Quốc tế |
|
| Quốc gia |
|
| Khác |
|
Từ khóa » Vòng đời Của Con Rắn Mối
-
Rắn Mối Thời Kì Sinh Sản
-
Rắn Mối Tự Nhiên Và Những Điều Bạn Cần Phải Biết
-
Hướng Dẫn Kỹ Thuật Nuôi Rắn Mối Hiệu Quả - UBND Tỉnh Cà Mau
-
Tìm Hiểu Kỹ Thuật Nuôi Rắn Mối Hiệu Quả Nhất
-
Hỏi- Đáp Về Rắn Mối
-
Một Số Tập Tính Của Rắn Mà Người Nuôi Cần Biết
-
Con Rắn Mối ăn Gì? Cách Nuôi? Giá Bao Nhiêu? Mua, Bán ở đâu?
-
Rắn Mối – Đặc điểm Sinh Học – Kỹ Thuật Nuôi – Đầu Ra
-
Kỹ Thuật Nuôi Rắn Mối | Kinh Nghiệm Làm ăn
-
Nuôi Rắn Mối - Báo Bình Dương Online
-
'Đời Sống ái ân' Kỳ Quặc ở Loài Rắn - BBC News Tiếng Việt
-
Vòng đời Của Nước, The Water Cycle, Vietnamese
-
Tuổi Thọ Của Rắn Là Bao Lâu? Đâu Là Loài Rắn Sống Lâu Nhất?